- Massospondylus
-
 Massospondylus
MassospondylusRango temporal: Jurásico inferior 
Representación artística de MassospondylusClasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Sauropodomorpha Infraorden: Prosauropoda (sin clasif.): Plateosauria Familia: Massospondylidae Género: Massospondylus Nombre binomial Massospondylus carinatus
Owen, 1854Sinonimia - Aetonyx Broom, 1911
- Aristosaurus Hoepen, 1920
- Dromicosaurus Hoepen, 1920
- Gryponyx Broom, 1912
- Gyposaurus Broom, 1911
- Hortalotarsus Seeley, 1894
- Leptospondylus Owen, 1895
- Pachyspondylus Owen, 1854
Massospondylus (gr. "vértebra elongada") es un género representado por una única especie de dinosaurio prosaurópodo masospondílido que vivió a principios del período Jurásico, entre 200 a 183 millones de años, desde el Hetangiano al Pliensbachiano en África. Descrita por Sir Richard Owen en 1854 a partir de restos descubiertos en Sudáfrica, siendo uno de los primeros dinosaurios en ser nombrado. Fósiles de este han sido encontrados en distintos lugares de Sudáfrica, Lesotho y Zimbabwe. Material adicional encontrado en la Formación Kayenta de Arizona , la India y Argentina han sido antiguamente asignados a este género, pero hoy se los considera distintos a Massospondylus.
Contenido
Descripción
Massospondylus fue un típico prosaurópodo de cuello medio largo y piernas delgadas.[1] Por un tiempo se creyó que Massospondylus era omnívoro pero ha habido nuevos descubrimientos que demuestran que era herbívoro. Este dato lo acerca más a sus descendientes. Para mejorar su digestion tragaba piedras (probado por la precencia de gastrolitos en su estómago) tal como lo hacían muchos sauropodomórfos. En cada una de sus patas delanteras poseía una garra aguda en el pulgar que fue utilizada en defensa o para recoger el alimento. Estudios recientes indican que Massospondylus crecía constantemente a través de su vida útil, que poseía sacos de aire similares a de los pájaros, y que pudo haber cuidado para sus crías.
Este dinosaurio alcanzaba alrededor de los 4 metros de largo,[2] [3] [4] y peso aproximadamente 135 kilogramos (,[4] aunque algunas estimacióes informan unos 6 metros de largo.[5] [6] Aunque siempre se ha asumido que habría sido cuadrúpedo, un estudio anatómico de sus miembros delanteros 2007 han cuestionado esto, sosteniendo que su rango de movimientos imposibilita un habitual paso cuadrúpedo eficaz. El estudio también eliminó la posibilidad del " caminar sobre los nudillos" y otras formas de locomoción que evitarían la aplicación de la capacidad limitada de Massospondylus a pronación de sus manos. Aunque su masa sugiera una naturaleza cuadrúpeda, realmente se trataría de un bipedo facultativo.[7]
 Massospondylus montado en el Museo de Historia Natural de Londres, mostrando una posición previa al estudio de 2007.
Massospondylus montado en el Museo de Historia Natural de Londres, mostrando una posición previa al estudio de 2007.
Massospondylus es un prosaurópodo típico en muchos otros aspectos. Poseía un cuerpo delgado con un cuello largo, con alrededor de nueve vértebras cervicales, trece dorsales, tres sacras y al menos 40 caudales. El pubis se prolongaba hacia adelante, como en la mayoría de los saurisquios. Tenía una estructura más ligera que la de Plateosaurus, que es visto como el típico prosarópodo.[8] Un reciente descubrimiento muestra que Massospondylus poseia un bien desarrollada clavícula, unida a una furcula, sugiriendo que ambos que sostuvieran inmóvil a la escapula y que las clavículas no eran rudimentarias y no funcionales en esos dinosaurios que no tenían espoletas verdaderas. Este descubrimiento también indica que la espoleta de los pájaro deriva de las clavículas.[9]
Como Plateosaurus, poseía cinco dedos en cada pie, con un largo pulgar para defenderse de los depredadores. Los cuartos y quintos dígitos de los patas delanteras eran minúsculos, dando a los brazos una posición ladeada. El estudio de 2007 indica que Massospondylus llevaba sus manos en una orientación semi supinación, con las superficies palmares haciendo frente a unas con otras, la muñeca nunca fueron encontradas giradas en especímenes fósiles articulados.[7]
Anatomia craneal
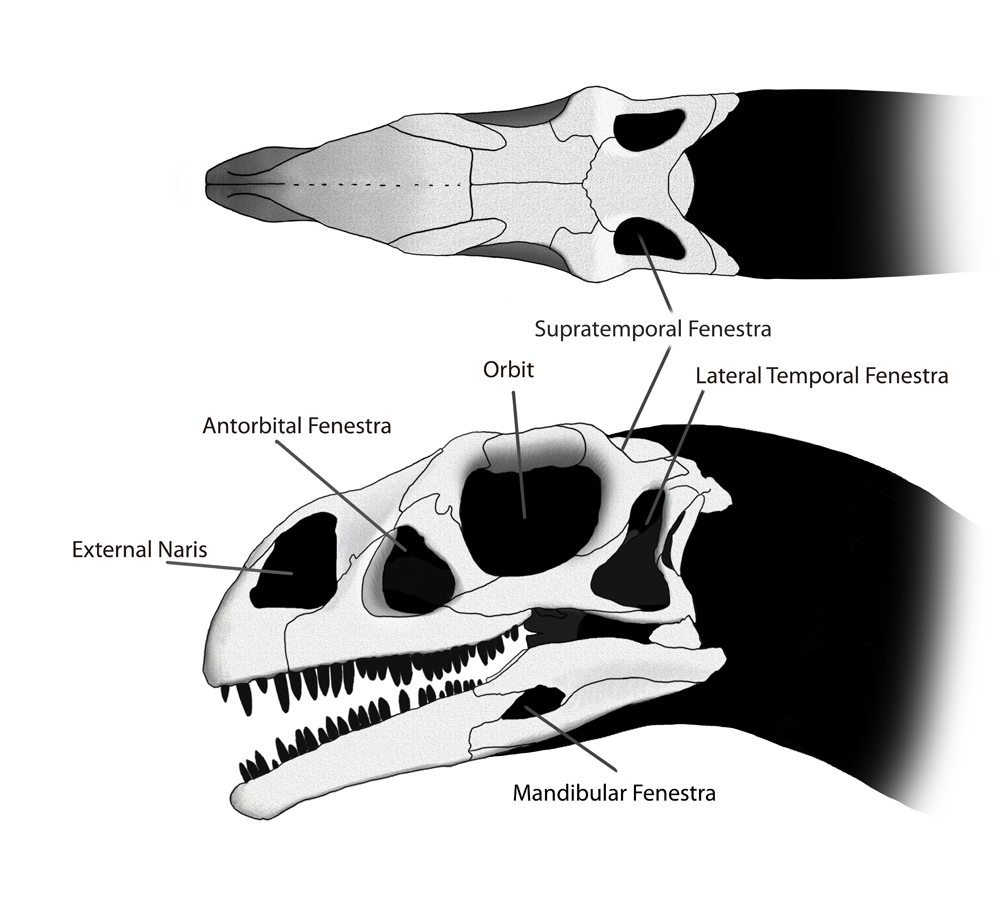
 Diagrama del cráneo de Massospondylus, mostrando varias de las aberturas craneales.
Diagrama del cráneo de Massospondylus, mostrando varias de las aberturas craneales.La pequeña cabeza de Massospondylus media aproximadamente la mitad del largo del fémur. Numerosos orificios, o fenestras, en el cráneo reducían el peso, suministrando espacio para la inserción de los músculos y los órganos de los sentidos. Estas fenestraciones se presentaban de a pares, una a cada lado del cráneo. En el frente del cráneo habían dos grandes y elípticas narinas. Las orbitas oculares eran proporcionalmente grandes en Massospondylus comparadas con otros géneros como Plateosaurus. La fenestra antorbital, era más pequeña que en Plateosaurus, estaba situada entre los ojos y la nariz. En la parte posterior del cráneo había dos pares más de fenestra temporal fenestra, la fenestra infratemporal e inmediatamente detrás de los cuencas de los ojo y de la fenestra supratemporale encima del cráneo. Una fenestra pequeña penetraba cada mandíbula.[8] La forma del cráneo se restaura tradicionalmente como más ancho y más corto que el de Plateosaurus, pero este aspecto puede ser debido aplastamiento experimentado por los varios especímenes.[8] Algunas características del cráneo son variables entre los individuos, por ejemplo, el grosor del borde superior de la órbita y de la altura trasera del maxilar. Estas diferencias pueden ser debido al dimorfismo sexual[10] o variación individual.[11]
Como en otros prosaurópodos, se ha propuesto que Massospondylus tuviese mejillas. Esta teoría fue propuesta porque hay algunos agujeros grandes para vasos sanguíneos en las superficies de los huesos de la quijada, a diferencia de los numerosos pequeños agujeros presentes en las quijadas de reptiles sin mejillas. Las mejillas habrían evitado que el alimento se les cayera de la boca cuando Massospondylus comía.[8] Crompton y Attridge (1986) describieron el cráneo de Massospondylus como poseedor de una maloclusión y sugiriendo la presencia de un pico córneo de queratina en la extremidad de la mandíbula inferior para suplir la diferencia en longitud y para explicar desgaste los dientes en la extremidad del hocico.[12] Sin embargo, posteriormente se ha demostrado que fue una interpretación basada un cráneo con la parte inferior aplastada dorsalmente. Los cráneos no aplastados en esta orientación no demuestran un maloclusión. [10] También parece haber una cierta variación de la morfología de l dientes basados en la posición de estos en la quijada.[13] La heterodoncia presente en Massospondylus es mayor que en Plateosaurus, aunque no es tan pronunciada como la especialización de los dientes en Heterodontosaurus.[13] Los dientes más cercano al frente del hocico tenían secciones redondeadas y eran afilado a los extremos, mientras que los dientes traseros, que eran espatulados y tenían secciones ovales.[8] [13]
Historia
Los primeros fósiles de Massospondylus fueron descritos por el paleontólogo Sir Richard Owen en 1854,[14] que le dio el nombre derivado de los términos del griego masson/μάσσων 'largo'[15] y spondylos/σπόνδυλος 'vértebra'.[16] Estos fósiles fueron encontrados en 1853 por J. M. Orpen en la Formación Elliot Superior en Harrismith, África del Sur. Estos restos constaban de las vértebras del cuello, del lomo, y cola, escapula, un húmero, una pelvis parcial, un fémur, una tibia y huesos de las manos y de los pies. El material holotipo original era parte de la colección de la universidad real Cirujanos en Londres que fue destruida durante Segunda Guerra Mundial, solamente los moldes se conservan.[17] Posibles restos de Massospondylus han sido encontrados en la Formación Elliot Superior, Formación Clarens y Arenisca Bushveld de Sudáfrica y Lesotho, la Arenisca Forest y Arenisca Karroo Superior de Zimbabwe y la Formación Kayenta de Arizona. Estos restos consisten en por lo menos 80 esqueletos parciales y cuatro cráneos, representando jóvenes y a adultos.[8]
Los reportes de Massospondylus de la Formación Kayenta enArizona esta basado en un cráneo encontrado en 1985. l cráneo encontrado en Kayenta es un 25% más largo que el espécimen más grande de África.[13] El especímen de Kayenta posee cuatro dientes en el premaxilar y dieciséis en el maxilar. Únicamente entre dinosaurios, también tenía minúsculos, de un milímetro, dientes palatales largos.[18] Recientes reestudios de los especímenes africanos de cráneos de Massospondylus, sin embargo, indica que no el espécimen de Kayenta no pertenece a Massospondylus.[19]
Massospondylus también ha sido reportado de Argentina,[8] pero recientemente ha sido reasignado a un género estrechamente vinculado. Los fósiles incluyeron varios esqueletos parciales y por lo menos un cráneo, encontrados en Formación Cañon del Colorado de la Provincia de San Juan, Argentina datada en el Jurásico Inferior. Este materia ha sido llamado Adeopapposaurus en 2009 por Ricardo N. Martínez.[20]
Especies
Se han nombrado muchas especies, aunque la mayoría no se consideren más válidas. M. carinatus, nombrado por Richard Owen, es la especie tipo.[21] Otras especie nombradas son M. browni (Seeley, 1895),[22] M. harriesi (Broom 1911),[23] M. hislopi (Lydekker, 1890),[24] M. huenei (Cooper, 1981),[25] M. rawesi (Lydekker, 1890),[24] and M. schwarzi (Haughton, 1924).[26]
M. browni, M. harriesi y M. schwarzi fueron todas encontradas en la Formación Elliot Superior Provincia del Cabo, Sudáfrica. Los tres se basan en el material fragmentario, y fueron considerados como dudosos en los más recientes trabajos.[8] M. browni se basa en dos vértebras del cuello, dos de la espalda y tres caudales y elementos varios de las patas.[22] M. harriesi por un miembro delantero.[23] M. schwarzi es conocido por un miembro tracero parcial y sacro.[26] M. hislopi y M. rawesi fueron nombrados por fósiles encontrados de la India.[24] M. hislopi se basa en vértebras del Triásico Superior en la Formación Maleri de Andhra Pradesh, considerando que M. rawesi esta basado en un diente del Cretácico Superior de la Formación Takli de Maharashtra.[27] M. hislopi fue tentativamente conservado como sauropodomorfo basal en las últimas revisiones,[8] pero M. rawesi es considerado un terópodo[28] o un no-dinosaurio.[17] M. huenei es una combinación creada por Cooper para Lufengosaurus huenei, cuando considero a Lufengosaurus y Massospondylus como sinónimos.[25] Esta sinónimia no fue mantenida por mucho tiempo.[8]
Nombre dudosos
A varios dinosaurios a menudo se los consideran sinónimos con Massospondylus. Estos incluyen a Aristosaurus, Dromicosaurus, Gryponyx, Hortalotarsus, Leptospondylus y Pachyspondylus, los cuáles son nombres dudosos de poco valor científico.[25] Hortalotarsus skirtopodus fue nombrado por Harry Seeley en 1894. Los fósiles consisten en los huesos parciales de la pierna. El año siguiente, Richard Owen nombró algunas vértebras fósiles Leptospondylus capensis y Pachyspondylus orpenii. Estos fósiles fragmentarios fueron destruidos en la Segunda Guerra Mundial. Aristosaurus erectus fue nombrado por E.C.N. van Hoepen en 1920 de acuerdo con un esqueleto casi completo. Hoepen también nombro a Dromicosaurus gracilis, el cuál consistió en un esqueleto parcial. Gryponyx taylori fue nombrado por Sidney H. Haughton en 1924, consistiendo en huesos de la cadera. Todos los fósiles antedichos vienen de las etapas fáunicas del Hetangiano o del Sinemuriano de Suráfrica, adonde Massospondylus ha sido encontrado.[29] [30] Bajo reglas de la nomenclatura zoológica, estos nombres son sinónimos menoress. Todos fueron nombrado después de que Massospondylus fuese descrito en un trabajo científico por lo que el nombre Massospondylus tiene la prioridad.
Clasificación
Massospondylus es un Prosauropoda, un agrupamiento de tempranos dinosaurios saurisquion que vivieron durante finales del Triásico a mediados del Jurásico, pero que había desaparecido para el finales de este ultimo período. Otros miembros del grupo incluyen Plateosaurus,[8] Yunnanosaurus,[8] y Riojasaurus.[31] La sistemática de los sauropodomórfos continúa experimentando una revisión, y muchos géneros que alguna vez se consideraron prosaurópodos clásicos se han quitado recientemente del grupo en la nomenclatura filogenética, considerando que su inclusión no constituiría un clado (el grupo natural que conteniene a todos los descendientes de un solo antepasado común). Exactamente que grupo constituye la monofilia de los prosarópodos no está claro. Yates yKitching (2003) publicaron un clado consistente de Riojasaurus, Plateosaurus, Coloradisaurus, Massospondylus, y Lufengosaurus.[32] Galton y Upchurch (2004) incluyeron a Ammosaurus, Anchisaurus, Azendohsaurus, Camelotia, Coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Mussaurus, Plateosaurus, Riojasaurus, Ruehleia, Saturnalia, Sellosaurus, Thecodontosaurus, Yimenosaurus y Yunnanosaurus en un monofiletico Prosauropoda.[8] Wilson (2005) considera a Massospondylus, Jingshanosaurus, Plateosaurus, y Lufengosaurus un grupo natural, con Blikanasaurus y Antetonitrus como posibles saurópodos.[33] Bonnan y Yates (2007) consideran a Camelotia, Blikanasaurus y Melanorosaurus posibles saurópodos.[34] Yates (2007) ubica Antetonitrus, Melanorosaurus, y Blikanasaurus como saurópodos basales y declina el uso del termino Prosauropoda, al que considera sinónimo de Plateosauridae. Sin embargo, él no eliminó la posibilidad de que un pequeño grupo de prosaurópodos que consistía en Plateosaurus , Riojasaurus , Massospondylus y sus parientes más cercanos fueran monofiléticos.[35]
Massospondylus es el género tipo para la propuesta familia Massospondylidae, a la que le da el nombre. La famila Massospondylidae también incluiría a Yunnanosaurus,[36] aunque Lu et al. (2007) ubican a Yunnanosaurusen su propia famila.[37] Yates (2007) considera a Massospondylus, Coloradisaurus y Lufengosaurus masospondílidos, con Yunnanosaurus en Anchisauria.[35] Smith y Pol (2007) también incluyeron a Massospondylidae en su [[análisis filogenético], incluyendo Massospondylus, Coloradisaurus y Lufengosaurus, así como su nuevo género, Glacialisaurus.[38] Adeopapposaurus, de acuerdo con los fósiles una vez considerado para pertenecer a una forma suramericana de Massospondylus, was also classified as un massospondílido.[20]
Filogenia
Plateosauria Plateosauridae Massopoda Riojasauridae (sin nombre) Massospondylidae Massospondylus
(sin nombre) (sin nombre) Filogenia de sauropodomorpha basal simplificado luego de Yates, 2007.[35] Aquí, Massospondylus es mostrado cercano a Lufengosaurus y Coloradisaurus. Esta es una sola de los posibles cladogramas de los sauropodomorfos basales. Paleoecología
La fauna y flora en el Jurásico temprano era similar en todo el planeta, coníferas adaptadas al clima calido, siendo las más comunes,[39] y los prosaurópodos y terópodos basales constituían la fauna mundial de dinosaurios.[40] El ambiente en el área del sudeste de África ha sido descrita como desértica.[41] Los Massospondylus africanos habrían sido contemporáneos a cocodrilomorfos, terápsidos tritilodontidos y tritelodóntidos mamiferos morganucodontidos,[8] y dinosaurios que incluian al terópodo Megapnosaurus rhodesiensis; una especie de Melanorosaurus (M. thabanensis); y varios géneros de ornitisquios tempranos como Lesothosaurus y los heterodontosáuridos Abrictosaurus, Heterodontosaurus y Lycorhinus.[30]
No esta claro que carnívoros habrían predado a Massospondylus. La mayor parte de los terópodos que se han descubierto en rocas jurásica temprana en África meridional, por ejemplo Megapnosaurus, eran más pequeños que los medianos Massospondylus. Estos depredadores más pequeños se creen que podrian realizar rapidos ataques para agotar los prosaurópodos, que habrían podido defenderse con sus grandes garras de la mano y pie.[41] El terópodo carnívoro Dracovenator de 6 metros de largo[42] vivió durante el mismo período que Massospondylus y también se ha encontrado en la Formación Elliot de Sudáfrica.[43]
Paleobiología
Como todo dinosaurio, mucho de la biología de Massospondylus, incluyendo su comportamiento, coloración, y fisiología, sigue siendo desconocidas. Sin embargo, los estudios recientes han permitido la especulación científica en temas tales como sus patrones de crecimiento,[44] dieta,[45] postura,[7] reproducción,[46] yrespiración.[47]
Un estudio de 2007 sugirió que Massospondylus puede haber utilizado sus brazos cortos en defensa de los, en combate intraspecificos, o en la alimentación, aunque sus brazos fueran demasiado cortos alcanzar su boca. Los científicos especulan que Massospondylus habría podido utilizar su gran garra del pulgar en combate, pelar el material vegetal de árboles ,[7] excavar, o para preparar nidos.[13]
Crecimiento
Un estudio de2005 indica que el taxón hermano de Massospondylus, Plateosaurus, exhibía patrones de crecimiento afectados por factores ambientales. El estudio indicó eso cuando el alimento era abundante, o cuando el clima era favorable, Plateosaurus exhibía un crecimiento acelerado. Estos patrones de crecimiento, llamados " plasticidad de desarrollo". Esto no es visto en otros dinosaurios, incluyendo Massospondylus, a pesar de la relación estrecha entre Plateosaurus y Massospondylus. El estudi indica que Massospondylus se desarrollo a lo largo de una trayectoria específica de crecimiento con poca variación en el índice de crecimiento y el tamaño final de un individuo.[44] Otro estudio indica que Massospondylus crecía a un máximo de 35 kilogramos por año y seguía creciendo al alcanzar los 15 años de edad.[48]
Dieta
Prosaurópodos como los masospondilos deben haber sido herbívoros u omnívoros. Tan recientemente como los años de 1980, paleontólogos discutían la posibilidad de carnivororismo en prosaurópodos.[25] [18] Sin embargo, la hipótesis de prosaurópodos carnívoros se ha desacreditado, y todos los estudios recientes favorecen una forma de vida herbívora u omnívora para estos animales. Galton y Upchurch (2004) encontraron que las características craneales (tales como articulación de la mandíbula) de la mayoría de los prosaurópodos están más cercano a las de reptiles herbívoros que los los carnívoros, y la forma de la corona dental es similar a las [[iguana]s herbívoras u omnívoras modernas. La anchura máxima de la corona era mayor que la de la raíz, dando por resultado un filo similar a los de reptiles herbívoros u omnívoros existentes.[8] Barrett (2000) ha propuesto que los prosaurópodos complementaran sus dietas herbívoras con presas pequeñas o carroña.[49] Gastrolitos (piedras estomacales) han sido encontradas en asociación con fósiles de masospondilos encontrados en Sudáfrica,[13] y en animal del tipo de los masospondilos del Triásico Superior en Virginia EE. UU..[45] Massospondylus tragó estas piedras para ayudar en la digestión,[13] con las contracciones musculares de la molleja que habría pulverizado el material vegetal tragado, compensando su incapacidad de masticar.
Reprodución
 Representación artística de la cría de Massospondylus, mostrada aquí como cuadrúpeda.
Representación artística de la cría de Massospondylus, mostrada aquí como cuadrúpeda.En 1977, siete huevos de fosilizados 190 millones de años Parqwue Nacional Golden Gate Highlands en Sudáfrica por James Kitching, quien los idenntifico como muy probablemente pertenecientes a masospondilos. Casi 30 años después de que la recolección de los fósiles se descubrieron en ellos embriones de 15 centímetros de largos. Estos siguen siendo los embriones más viejos de dinosaurio jamas encontrados. Notablemente, los más desarrollados no tenian ningun diente, sugiriendo ellos no tenían ninguna manera de alimentarse. Debido a la carencia de dientes y las proporciones del cuerpo, los científicos especulan que el cuidado postnatal pudo haber sido necesario. Las cuatro piernas de los embriones tenian una longitud igual, indicando que las crias de masospondilos erán cuadrúpedas.[46] El cráneo y los ojos eran proporcionalmente de gran tamaño comparado a los adultos, cosa común en otros vertebrados muy jóvenes, aunque los cráneos de los jóvenes fueran más altos y estrechos.[13] La posición cuadrupeda de las crías sugiere que la postura cuadrúpeda del los saurópodos adultos pudo haberse desarrollado más adelante por la retención de características juveniles, un fenómeno evolutivo conocido como pedomorfosis. [46]
Sistema respiratorio
Muchos dinosaurios saurisquios poseen vértebras y costillas repletas de cavidades para el aire, forámenes neumáticos, lo cuál redujo el peso de los huesos y puede haber servido como un básico sistema de ventilación por flujo similar a el de pájaros modernos. En tal sistema, las vértebras y las costillas del cuello son ahuecadas por el saco de aire cervical, las vértebras de la espalda, por el pulmón y el lomo y vértebras sacras, por el saco de aire abdominal. Estos órganos constituyen un complejo y un método muy eficiente de respiración.[50] Los prosaurópodos son el único grupo principal de saurisquios sin un sistema extenso de foramenes neumáticos. Aunque sea posible que las muescas neumáticas se hayan encontrado en Plateosaurus y Thecodontosaurus, aunque eran muescas muy pequeñas. Un estudio en 2007 concluyó que los prosaurópodos como Massospondylus tuvo probablemente sacos de aire abdominal y cervical, basandoce en la evidencia de ellos y en taxones hermanos (los terópodos y los saurópodos). El estudio concluyó que era imposible determinar si los prosaurópodos tenían un sistema de ventilación de flujo, como los pájaros, pero casi ciertamente los sacos de aire estaban presentes.[47]
Referencias
- ↑ Senter, P. & Bonnan, M. 2005. Evidence for obligate bipedality in the basal sauropodomorphs Plateosaurus and Massospondylus. Journal of Vertebrate Paleontology. Abstracts of papers. Sixty-fifth annual meeting Society of vertebrate paleontology, Mesa Southwest Museum and Phoenix Marriott Mesa, Mesa, Arizona. October 19-22, 25, Supplement to number 3, 114A.
- ↑ Lambert, David; and the Diagram Group (1983). A Field Guide to Dinosaurs. New York: Avon Books. p. 103. ISBN 0-380-83519-3.
- ↑ Lambert, David; and the Diagram Group (1990). The Dinosaur Data Book. New York: Avon Books. p. 75. ISBN 0-380-75896-3.
- ↑ a b Seebacher, Frank (2001). «A new method to calculate allometric length-mass relationships of dinosaurs». Journal of Vertebrate Paleontology 21 (1): pp. 51–60. doi:.
- ↑ Lessem, Don; Donald F. Glut (1993). «Massospondylus». The Dinosaur Society Dinosaur Encyclopedia. Random House. p. 295. ISBN 0-679-41770-2.
- ↑ Glut, Donald F. (2000). «Massospondylus». Dinosaurs: The Encyclopedia: Supplement One. Jefferson, North Carolina: McFarland & Co. p. 258. ISBN 0-7864-0591-0.
- ↑ a b c d ,, Matthew; Phil Senter (2007). «Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds?». En Paul M. Barrett & D. J. Batten (eds.). Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs. Special Papers in Palaeontology 77. London: The Palaeontological Association. pp. 139–155. ISBN 978-1-4051-6933-2.
- ↑ a b c d e f g h i j k l m n ñ Galton, P.M. and Upchurch, P. (2004). "Prosauropoda". Weishampel & als: The Dinosauria (2nd edition), pp. 232–258.
- ↑ Yates, Adam M.; and Vasconcelos, Cecilio C. (2005). «Furcula-like clavicles in the prosauropod dinosaur Massospondylus». Journal of Vertebrate Paleontology 25 (2): pp. 466–468. doi:.
- ↑ a b Gow, Christoper E.; J.W. Kitching and Michael K. Raath (1990). «Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research». Palaeontologia Africana 27: pp. 45–58.
- ↑ Galton, Peter M. (1997). «Comments on sexual dimorphism in the prosauropod dinosaur Plateosaurus engelhardti (Upper Triassic, Trossingen)». Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 11: pp. 674–682.
- ↑ Crompton, A.W.; John Attridge (1986). «Masticatory apparatus of the larger herbivores during Late Triassic and Early Jurassic times». En Kevin Padian (ed.). The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary. New York: Cambridge University Press. pp. 223–236. ISBN 0-521-36779-4.
- ↑ a b c d e f g h "Massospondylus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 39. ISBN 0-7853-0443-6.
- ↑ Owen, Richard (1854). "Descriptive catalogue of the Fossil organic remains of Reptilia and Pisces contained in the Museum of the Royal College of Surgeons of England." London p. 1–184 OCLC 14825172
- ↑ Creisler, Ben (2003). «Dinosauria Translation and Pronunciation Guide». Dinosauria.com. Consultado el 23-11-2007.
- ↑ Liddell, Henry George; Robert Scott (1980). A Greek-English Lexicon (Abridged Edition). Oxford: Oxford University Press. ISBN 0-19-910207-4. OCLC 17396377.
- ↑ a b Glut, Donald F. (1997). «Massospondylus». Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 581–586. ISBN 0-89950-917-7.
- ↑ a b Attridge, J.; A.W. Crompton and Farish A. Jenkins, Jr. (1985). «The southern Liassic prosauropod Massospondylus discovered in North America». Journal of Vertebrate Paleontology 5 (2): pp. 128–132.
- ↑ Sues, H.-D.; R.R. Reisz, S. Hinic and M.A. Raath (2004). «On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens formations (Lower Jurassic) of South Africa». Annals of Carnegie Museum 73 (4): pp. 239–257.
- ↑ a b Martínez, Ricardo N. (2009). «Adeopapposaurus mognai, gen. et sp. nov (Dinosauria: Sauropodomorpha), with comments on adaptations of basal sauropodomorpha». Journal of Vertebrate Paleontology 29 (1): pp. 142–164.
- ↑ «Massospondylus». The Paleobiology Database (2007). Consultado el 03-11-2007.
- ↑ a b Seeley, H.G. (1895). «On the type of the genus Massospondylus and on some Vertebrae and limb-bone of M. (?) browni». Annals and Magazine of Natural History 15: pp. 102–125.
- ↑ a b Broom, Robert (1911). «On the dinosaurs of the Stormberg, South Africa». Annals of the South African Museum 7 (4): pp. 291–308.
- ↑ a b c Lydekker, Richard (1890). «Note on certain vertebrate remains from the Nagpur district». Records of the Geological Survey of India 23 (1): pp. 21–24.
- ↑ a b c d Cooper, M.R. (1980). «The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance». Occasional Papers of the National Museums and Monuments of Rhodesia, Series B, Natural Sciences 6 (10): pp. 689–840.
- ↑ a b Haughton, Sydney H. (1924). «The fauna and stratigraphy of the Stormberg Series». Annals of the South African Museum 12: pp. 323–497.
- ↑ Weishampel, David B. (1990). «Dinosaur distribution». En David B. Weishampel, Peter Dodson and Halszka Osmólska (eds.). The Dinosauria (1st ed. edición). Berkeley: University of California Press. pp. 63–139. ISBN 0-520-06727-4.
- ↑ Molnar, R.E. (1990). Problematic Theropoda: "Carnosaurs". In Weishampel & als: The Dinosauria (1st ed.), pp. 306–317.
- ↑ Tweet, Justin (2007). «Sauropodomorpha». Thescelosaurus!. Consultado el 16-12-2007.
- ↑ a b Weishampel, David B.; Paul M. Barrett, Rodolfo Coria, A., Jean Le Loeuff, Zhao Xijin Xu Xing, Ashok Sahni, Elizabeth M.P. Gomani and Christopher R. Noto (2004). «Dinosaur Distribution». En David B. Weishampel, Peter Dodson and Halszka Osmólska (eds.). The Dinosauria (2nd ed. edición). Berkeley: University of California Press. pp. 517–606. ISBN 0-520-24209-2.
- ↑ Galton, P.M (2005). «Postcranial anatomy of referred specimens of the sauropodomorph dinosaur Melanorosaurus from the Upper Triassic of South Africa». En Virginia Tidwell and Kenneth Carpenter (eds.). Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington: Indiana University Press. pp. 1–37. ISBN 0-253-34542-1.
- ↑ Yates, Adam M.; James W. Kitching (2003). «The earliest known sauropod dinosaur and the first steps towards sauropod locomotion». Proceedings of the Royal Society B 270 (1525): pp. 1753–1758. doi:.
- ↑ Wilson, Jeffrey A.; Curry Rogers, Kristina (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 15–49. ISBN 978-0-520-24623-2.
- ↑ Bonnan, Matthew F. and Adam M. Yates, "A new description of the forelimbs of the basal sauropodomorph Melanorosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 157–168.
- ↑ a b c Yates, Adam M., "The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria)". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 9–55.
- ↑ Sereno, Paul C. (1999). «The evolution of dinosaurs». Science 284 (5423): pp. 2137–2147. doi:. PMID 10381873.
- ↑ Lu, J.; T. Li, S. Zhong, Y. Azuma, M. Fujita, Z Dong and Q. Ji (2007). «New yunnanosaurid dinosaur (Dinosauria, Prosauropoda) From the Middle Jurassic Zhanghe Formation of Yuanmou, Yunnan province of China». Memoir of the Fukui Prefectural Dinosaur Museum 6: pp. 1–15.
- ↑ Smith, Nathan D.; Diego Pol (2007). «Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica» (pdf). Acta Palaeontologica Polonica 52 (4): pp. 657–674. http://www.app.pan.pl/acta52/app52-657.pdf.
- ↑ Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. p. 45. ISBN 1-55971-038-1.
- ↑ Holtz, Thomas R., Jr., Ralph E. Chapman and Matthew C. Lamanna, "Mesozoic biogeography of Dinosauria". In Weishampel & als., The Dinosauria (2nd edition), pp. 627–642.
- ↑ a b Paul, Gregory S. (1988). «Coelophysis rhodesiensis». Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 262–265. ISBN 978-0-6716-1946-6.
- ↑ Smith, N.D.; P.J. Makovicky, D. Pol, W.R. Hammer and P.J. Currie (2007). «The Dinosaurs of the Early Jurassic Hanson Formation of the Central Transantarctic Mountains: Phylogenetic Review and Synthesis». En Alan Cooper, Carol Raymond et als. (eds.) (PDF). Antarctica: A Keystone in a Changing World—Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences. Reston, Virginia: U.S. Geological Survey. doi:. ISBN 1-411-31788-2. OCLC 85782509. http://pubs.usgs.gov/of/2007/1047/srp/srp003/of2007-1047srp003.pdf.
- ↑ Yates, A.M. (2006). «A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods». Palaeontologia africana 41: pp. 105–122.
- ↑ a b Sander, P. Martin; Nicole Klein (2005). «Developmental plasticity in the life history of a prosauropod dinosaur». Science 310 (5755): pp. 1800–1802. doi:. PMID 16357257.
- ↑ a b Weems, Robert E.; Michelle J. Culp and Oliver Wings (2007). «Evidence for Prosauropod Dinosaur Gastroliths in the Bull Run Formation (Upper Triassic, Norian) of Virginia». Ichnos 13 (3–4): pp. 271–295. doi:.
- ↑ a b c Reisz, Robert R.; Diane Scott, Hans-Dieter Sues, David C. Evans, Michael A. Raath (2005). «Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance». Science 309 (5735): pp. 761–764. doi:. PMID 16051793.
- ↑ a b Wedel, Mathew (2007). «What pneumaticity tells us about 'prosauropods', and vice versa» (PDF). Special Papers in Palaeontology 77: pp. 207–222. http://sauroposeidon.net/Wedel_2007_pneumaticity-and-prosauropods.pdf.
- ↑ Erickson, Gregory M.; Kristina Curry Rogers and Scott A. Yerby (2001). «Dinosaurian growth patterns and rapid avian growth rates». Nature 412: pp. 429–433. doi:.
- ↑ Barrett, P.M. (2000). «Prosauropod dinosaurs and iguanas: Speculations on the diets of extinct reptiles». En Hans-Dieter Sues. Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record. Cambridge: Cambridge University Press. pp. 42–78. ISBN 978-0-521-59449-3.
- ↑ O'Connor, Patrick M.; Leon P.A.M. Claessens (2006). «Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs». Nature 436 (7048): pp. 253–256. doi:.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre MassospondylusCommons.
Wikimedia Commons alberga contenido multimedia sobre MassospondylusCommons. Wikiespecies tiene un artículo sobre Massospondylus. Wikispecies
Wikiespecies tiene un artículo sobre Massospondylus. Wikispecies- Locomotion and Ontogeny of Massospondylus carinatus de la Universidad de Toronto en Missisauga.
- Massospondylidae de Palaeos.com (técnica).
- Massospondylus en DinoData.org.
- Massospondylus de PrimeOrigins.co.za (para niños).
- "Oldest known dinosaur embryos identified" Univesidad de Witwatersrand.
- Sauropodomorpha en Thescelosaurus!
Categorías:- Fósiles
- Prosauropoda
- Dinosaurios del Triásico
- Dinosaurios norteamericanos
- Dinosaurios africanos


Wikimedia foundation. 2010.