- Transporte de membrana
-
En biología celular se denomina transporte de membrana al conjunto de mecanismos que regulan el paso de solutos, como iones y pequeñas moléculas, a través de membranas plasmáticas, esto es, bicapas lipídicas que poseen proteínas embebidas en ellas. Dicha propiedad se debe a la selectividad de membrana, una característica de las membranas celulares que las faculta como agentes de separación específica de sustancias de distinta índole química; es decir, la posibilidad de permitir la permeabilidad de ciertas sustancias pero no de otras.[1]
Los movimientos de casi todos los solutos a través de la membrana están mediados por proteínas transportadoras de membrana, más o menos especializadas en el transporte de moléculas concretas. Puesto que la diversidad y fisiología de las distintas células de un organismo está relacionada en buena medida con su capacidad de captar unos u otros elementos externos, se postula que debe existir un acervo de proteínas transportadoras específico para cada tipo celular y para cada momento fisiológico determinado;[1] dicha expresión diferencial se encuentra regulada mediante: la transcripción diferencial de los genes codificantes para esas proteínas y su traducción, es decir, mediante los mecanismos genético-moleculares, pero también a nivel de la biología celular: dichas proteínas pueden requerir de activación mediada por rutas de señalización celular, activación a nivel bioquímico o, incluso, de localización en vesículas del citoplasma.[2]
Contenido
Generalidades
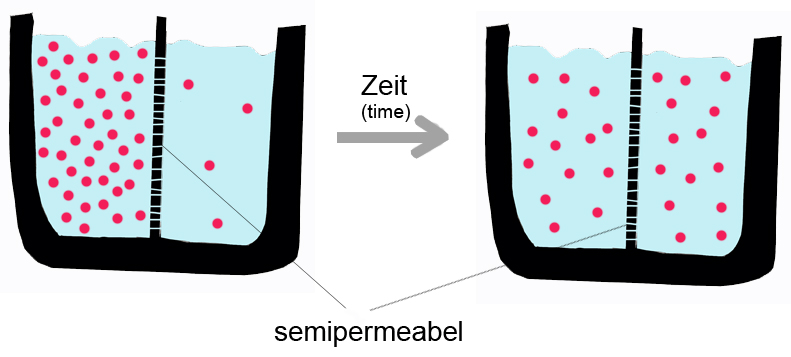
Termodinámicamente, el flujo de sustancias de un compartimento a otro puede realizarse a favor o en contra de un gradiente, ya sea de concentración, o electroquímico. Si el intercambio de sustancias se realiza a favor del gradiente, esto es, en el sentido de los potenciales decrecientes, el requerimiento de energía externo al sistema es nulo; si, en cambio, el transporte se hace en contra del gradiente, se requiere el aporte de energía, energía metabólica en nuestro caso.[3] Por ejemplo, un mecanismo químico de separación clásico que no requiere un aporte de energía externo es la diálisis: en ella, una membrana semipermeable separa dos soluciones que difieren en la concentración de un mismo soluto. Si la membrana permite el paso de agua pero no el del soluto, sucede que el agua fluye hacia el compartimento más concentrado en soluto, a fin de establecer un equilibrio en el cual la energía del sistema sea mínima. Para que suceda este flujo, puesto que el agua se desplaza de un lugar muy concentrado a uno muy diluido en disolvente (en cuanto a soluto, se da la situación opuesta), y, por ello, lo hace a favor de gradiente, no se requiere un aporte de energía externo.
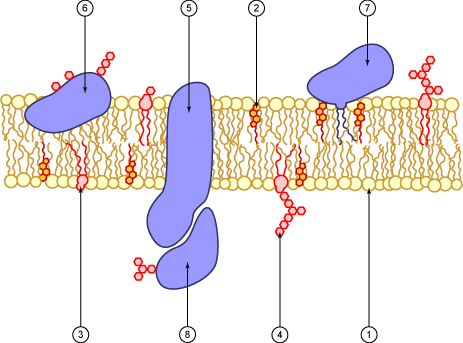
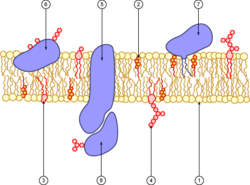 Diagrama de una membrana plasmática. 1. fosfolípido 2. colesterol 3. glicolípido 4. glúcido 5. proteína transmembrana 6. glicoproteína 7. proteína anclada a un fosfolípido 8. glicoproteína periférica.
Diagrama de una membrana plasmática. 1. fosfolípido 2. colesterol 3. glicolípido 4. glúcido 5. proteína transmembrana 6. glicoproteína 7. proteína anclada a un fosfolípido 8. glicoproteína periférica.
La naturaleza de las membranas biológicas, especialmente la de sus lípidos, es anfipática, lo que se traduce en que forman una bicapa que alberga una parte interna hidrofóbica y una externa hidrofílica, permite que surja una posibilidad de transporte, la difusión simple o difusión pasiva, que consiste en la difusión de sustancias a su través sin gasto de energía metabólica y sin ayuda de proteínas transportadoras. En el caso de que la sustancia a transportar posea una carga neta, difundirá no sólo en respuesta a un gradiente de concentración, sino también al potencial de membrana, esto es, al gradiente electroquímico.
- Permeabilidad relativa de una bicapa de fosfolípidos a distintas sustancias[1]
Tipo de sustancia Ejemplos Comportamiento Gases CO2, N2, O2 Permean Moléculas polares pequeñas sin carga Urea, agua, etanol Permean, total o parcialmente Grandes moléculas polares sin carga glucosa, fructosa No permean Iones K+, Na+, Cl-, HCO3- No permean Moléculas polares cargadas ATP, aminoácidos, glucosa-6-fosfato No permean Puesto que son pocas las moléculas que son capaces de difundir a través de una membrana lipídica, la mayoría de los procesos de transporte involucran a proteínas de transporte. Se trata de proteínas transmembrana que poseen multitud de hélices alfa inmersas en la matriz lipídica o bien, al menos en bacterias, beta láminas.[4] Dicha estructura probablemente implique una vía de entrada a través de entornos hidrofílicos proteicos que causarían una disrupción en el medio altamente hidrofóbico constituido por los lípidos.[1] Las proteínas intervienen de diversas formas en el transporte: actúan tanto como bombas impulsadas por ATP, esto es, por energía metabólica, o como canales de difusión facilitada.
Termodinámica
Un proceso fisiológico sólo puede llevarse a cabo si no contraviene los principios termodinámicos elementales. El transporte de membrana obedece algunas leyes físicas que definen sus capacidades y por ello su utilidad biológica.
Un principio general de la termodinámica que gobierna la transferencia de sustancias a través de membranas u otras superficies es que el cambio de la energía libre, ΔG, para el transporte de un mol de una sustancia concentrada a C1 en un compartimento, hacia un lugar en el que esté a C2, es de:[5]
Por lo que si C2 es menor que C1, ΔG es también negativo, y el proceso es termodinámicamente favorable. Conforme se va transfiriendo la energía de un compartimento a otro, y salvo que intervengan otros factores, se llega a un equilibrio donde C2=C1, y por ello ΔG=0. No obstante, existen tres circunstancias en las que puede evitarse esta igualdad, circunstancias vitales para el desempeño de la función in vivo de las membranas biológicas:[5]
- Las macromoléculas de un lado de la membrana pueden unir específicamente a un determinado compuesto o modificarlo químicamente. De este modo, aunque la concentración del compuesto sea realmente diferente a ambos lados de la membrana, su disponibilidad reducida en uno de los compartimentos puede hacer que, a efectos prácticos, no exista un gradiente que favorezca el transporte.
- Puede existir un potencial eléctrico de membrana a través de ella que influya en la distribución de iones. Por ejemplo, para un proceso en el que el transporte de iones se de desde el exterior al interior, sucede que:
- Donde F es la constante de Faraday y ΔP el potencial de membrana, en voltios. Si ΔP es negativo y Z es positivo, el término ZFΔP contribuye de forma negativa a ΔG, esto es, favorece el transporte de cationes hacia el interior de la célula. Esto es, si la diferencia de potencial se mantiene, el estado de equilibrio ΔG=0 no corresponderá a una cantidad equimolecular de iones a ambos lados de la membrana.
- Si se acopla un proceso con una ΔG negativa al proceso, el ΔG global deberá ser modificado. Esta situación, común en el transporte activo, se describe según:
- Donde ΔGb corresponde a una reacción favorecida termodinámicamente, como la hidrólisis de ATP, o el cotransporte de un compuesto que se desplace a favor de su gradiente.
Tipos
Difusión simple
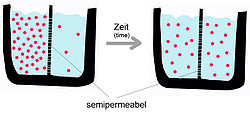 Una membrana semipermeable separa dos compartimentos con concentraciones distintas de un soluto: con el paso del tiempo, el soluto difundirá hasta alcanzar el equilibrio a ambos lados.
Una membrana semipermeable separa dos compartimentos con concentraciones distintas de un soluto: con el paso del tiempo, el soluto difundirá hasta alcanzar el equilibrio a ambos lados.Como se mencionó anteriormente, la difusión pasiva es un fenómeno espontáneo puesto que suceden incrementando la entropía del sistema, y disminuyendo la energía libre.[5] No requiere de la intervención de proteínas de membrana, pero sí de las características de la sustancia a transportar y de la naturaleza de la bicapa. Para el caso de una membrana fosfolipídica pura, la velocidad de difusión de una sustancia depende de su:
- gradiente de concentración,
- hidrofobicidad,
- tamaño,
- carga, si la molécula posee carga neta.
Estos factores afectan de diversa manera a la velocidad de difusión pasiva:
- a mayor gradiente de concentración, mayor velocidad de difusión,
- a mayor hidrofobicidad, esto es, mayor coeficiente de partición, mayor solubilidad en lípido y por tanto mayor velocidad de difusión,
- a mayor tamaño, menor velocidad de difusión,
- dado un potencial de membrana, es decir, la diferencia de potencial entre la cara exoplasmática y la endoplasmática de la membrana, y un gradiente de concentración se define un gradiente electroquímico que determina las direcciones de transporte energéticamente favorables de una molécula cargada, dependiendo de la naturaleza de ésta y del signo del potencial, si bien la mayor parte de las células animales poseen carga negativa en su exterior.[6]
La difusión simple a través de la membrana lipídica muestra una cinética de no saturación, esto es, que, puesto que la tasa neta de entrada está determinada sólo por la diferencia en el número de moléculas a cada lado de la membrana, la entrada aumenta en proporción a la concentración de soluto en el fluido extracelular. Esta característica distingue la difusión simple de los mecanismos de penetración por canales de transporte mediado.[6]
Difusión facilitada
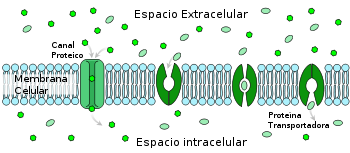 La difusión facilitada involucra el uso de un proteína para facilitar el movimiento de moléculas a través de la membrana. En algunos casos, las moléculas pasan a través de canales con la proteína. En otros casos, la proteína cambia su forma, permitiendo que las moléculas pasen a través de ella.
La difusión facilitada involucra el uso de un proteína para facilitar el movimiento de moléculas a través de la membrana. En algunos casos, las moléculas pasan a través de canales con la proteína. En otros casos, la proteína cambia su forma, permitiendo que las moléculas pasen a través de ella.Bajo el mismo principio termodinámico que en el caso de la difusión simple, es decir, que el soluto a transportar lo hace a favor de gradiente, la difusión facilitada opera de modo similar, pero está facilitada por la existencia de proteínas canal, que son las que facilitan el transporte de, en este caso, agua o algunos iones y moléculas hidrófilas. Estas proteínas integrales de membrana conforman estructuras en forma de poro inmersas en la bicapa, que dejan un canal interno hidrofílico que permite el paso de moléculas altamente lipófobas como las mencionadas anteriormente. La apertura de este canal interno puede ser constitutiva, es decir, continua y desregulada, en los canales no regulados, o bien puede requerir una señal que medie su apertura o cierre: es el caso de los canales regulados.[1]
Transporte activo y cotransporte
En él se efectúa un transporte en contra del gradiente de concentración o electroquímico y, para ello, las proteínas transportadoras implicadas consumen energía metabólica (comúnmente adenosín trifosfato). La hidrólisis del compuesto que actúa como moneda energética puede ser muy evidente, como en el caso de los transportadores que son ATPasas, o puede tener un origen indirecto: por ejemplo, los cotransportadores emplean gradientes de determinados solutos para impulsar el transporte de un determinado compuesto en contra de su gradiente, a costa de la disipación del primer gradiente mencionado. Pudiera parecer que en este caso no interviene un gasto energético, pero no es así puesto que el establecimiento del gradiente de la sustancia transportada colateralmente al compuesto objetivo ha requerido de la hidrólisis de ATP en su generación mediante unos determinados tipos de proteínas denominados bombas.[2] Por ello, se define transporte activo primario como aquél que hidroliza ATP de forma directa para transportar el compuesto en cuestión, y transporte activo secundario como aquél que utiliza la energía almacenada en un gradiente electroquímico.
El descubrimiento de la existencia de este tipo de transportadores se produjo al estudiar cinéticamente la transferencia de moléculas a través de las membranas: para algunos solutos, se observó que la velocidad de entrada alcanza una meseta a partir de cierta concentración externa a partir de la cual no se produce un incremento significativo de velocidad de captación, esto es, surge una respuesta tipo curva logística. Se interpretó que el transporte aquí se produce por la formación de un complejo sustrato-transportador, conceptualmente idéntico al complejo enzima-sustrato de la cinética enzimática. Por ello, cada proteína transportadora posee una constante de afinidad por el soluto que es igual a la concentración del soluto cuando la velocidad de transporte es la mitad de su valor máximo (equivaldría, para el caso de un enzima, a la constante de Michaelis-Menten).[6]
Algunos rasgos importantes del transporte activo, además de su capacidad de intervenir aun en contra de gradiente, su cinética y el empleo de ATP, son: su elevado grado de selectividad y su facilidad de inhibición farmacológica selectiva.[6]
Transportadores
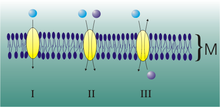 Uniporte, simporte y antiporte de moléculas a través de transportadores de membrana.
Uniporte, simporte y antiporte de moléculas a través de transportadores de membrana.Un transportador puede movilizar diversos iones y moléculas. Según la direccionalidad, se distinguen:
- antiportadores: aquellos que transportan un tipo de molécula en contra de su gradiente al mismo tiempo que desplazan uno o más iones diferentes a favor del suyo, siendo ambos gradientes contrapuestos.
- simportadores: los que desplazan el compuesto a transportar en contra de su gradiente acoplando este trasiego al desplazamiento de uno o más iones diferentes a favor del suyo, que, en este caso, es equivalente al de la molécula a transportar.
Ambos reciben el nombre de cotransportadores.
Bombas
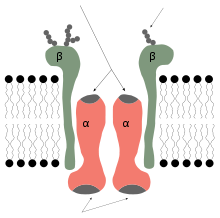 Diagrama simplificado de una bomba de sodio.
Diagrama simplificado de una bomba de sodio.Una bomba es una proteína que hidroliza ATP para transportar a través de una membrana un determinado soluto a fin de generar un gradiente electroquímico que confiera unas características de potencial a ésta. Dicho gradiente posee un interés por sí mismo para la definición del estado de la célula, como es el potencial de Nernst, pero interviene activamente en el transporte de sustancias a través de la membrana, que es el tema aquí tratado, puesto que aporta un aumento de entropía al sistema en caso de cotransporte de sustancias que se encuentran en trasiego en contra de su gradiente.
Una de las bombas de mayor relevancia en células animales es la bomba sodio-potasio, que opera mediante el mecanismo siguiente:[7]
- Unión de tres Na+ a sus sitios activos.
- Fosforilación de la cara citoplasmática de la bomba que induce a un cambio de conformación en la proteína. Esta fosforilación se produce por la transferencia del grupo terminal del ATP a un residuo de ácido aspártico de la proteína.
- El cambio de conformación hace que el Na+ sea liberado al exterior.
- Una vez liberado el Na+, se unen dos moléculas de K+ a sus respectivos sitios de unión de la cara extracelular de la proteína.
- La proteína se desfosforila produciéndose un cambio conformacional de esta, lo que produce una transferencia de los iones de K+ al citosol.
Selectividad de membrana
Puesto que la característica primordial del transporte a través de una membrana biológica es la selectividad de ésta y su subsiguiente actuación como barrera específica para determinadas sustancias, la fisiología subyacente de este fenómeno ha sido estudiada profusamente. Clásicamente se ha dividido el estudio de esta propiedad en lo referente a electrolitos y a no electrolitos.
Selectividad para electrolitos
Los canales iónicos definen un diámetro interno que permite el paso de pequeños iones de forma más o menos específica. Puesto que el tamaño del ion está relacionado con la especie química, se podría asumir a priori que un canal cuyo diámetro de poro fuera suficiente para el paso de un ion permitiría asimismo el trasiego de otros de menor tamaño, cosa que no sucede en la mayoría de los casos. Existen dos características ajenas al tamaño que son importantes en la determinación de la selectividad de los poros de la membrana: la facilidad de deshidratación e interacción con las cargas del interior del poro.[6]
Para que un ion penetre en el poro, debe disociarse de las moléculas de agua que lo recubren en sucesivas capas de solvatación. La tendencia a deshidratarse, o la facilidad para hacerlo, está relacionada con el tamaño del ion: los iones grandes lo hacen con más facilidad que los pequeños, por lo que un poro con centros polares débiles admitirá preferentemente iones grandes, antes que pequeños.[6]
Cuando el interior del canal está tapizado de grupos polares, procedentes de las cadenas laterales de sus aminoácidos componentes,[7] la interacción del ion deshidratado con estos centros puede ser más importante que la facilidad de deshidratación para conferir la especificidad del canal. Por ejemplo, un canal revestido de histidinas y argininas, con grupos cargados positivamente, repelerá de forma selectiva a los iones cargados con el mismo signo, pero facilitará el paso de los cargados negativamente. Además, en este caso, los iones más pequeños pueden interactuar de forma más cercana por cuestiones estéricas, lo cual incrementa en mucho las interacciones carga-carga y, por tanto, exagera el efecto.[6]
Selectividad para no electrolitos
Los no electrolitos, sustancias que generalmente son hidrofóbicas y lipofílicas, suelen atravesar la membrana por disolución en la bicapa lipídica y, por tanto, mediante difusión simple. La facilidad para difundir en este caso es dependiente del coeficiente de partición K, por lo general, si bien existen algunos no electrolitos que atraviesan la membrana por transporte mediado por un transportador.
En el caso de que el no electrolito esté parcialmente cargado, es decir, sea más o menos polar, como es el caso del etanol, metanol o urea, se permite el paso a través de la membrana mediante canales acuosos inmersos en la membrana. Es interesante recalcar que no existe un mecanismo de regulación efectivo que establezca barreras a este transporte, lo que implica una vulnerabilidad intrínseca de las células a la penetración de estas moléculas.[6]
Véase también
Referencias
- ↑ a b c d e Lodish et al. (2005). Biología celular y molecular. Buenos Aires: Médica Panamericana. ISBN 950-06-1974-3.
- ↑ a b Alberts et al (2004). Biología molecular de la célula. Barcelona: Omega. ISBN 54-282-1351-8.
- ↑ Cromer, A.H. (1996). Física para ciencias de la vida. Reverté ediciones. ISBN para España 84-291-1808-X.
- ↑ Prescott, L.M. (1999). Microbiología. McGraw-Hill Interamericana de España, S.A.U.. ISBN 84-486-0261-7.
- ↑ a b c Mathews, C. K.; Van Holde, K.E et Ahern, K.G (2003). Bioquímica (3ª edición). ISBN 84-7892-053-2.
- ↑ a b c d e f g h Randall, D.; Burggren, W. et French, K. (1998). Eckert Fisiología animal (4ª edición). ISBN 84-486-0200-5.
- ↑ a b Lehninger, Albert (1993). Principles of Biochemistry, 2nd Ed.. Worth Publishers. ISBN 0-87901-711-2.
Categorías:- Fisiología celular
- Biología molecular





Wikimedia foundation. 2010.