- Tubulina
-
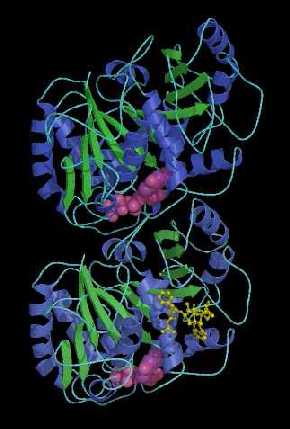
 Diagrama de cintas basado en un modelo de una resolución de 3.7 Angstrom desarrollado por científicos del laboratorio de Berkeley, que muestra que la tubulina es un heterodímero que consiste de dos monómeros de estructuras casi idénticas (alfa y beta). Cada monómero está formado por un núcleo de dos láminas beta (verde), rodeado por hélices (azul), y cada uno se une a un nucleótido guanina (rosa). Además de un sitio de unión a un nucleótido, cada monómero tiene otros dos sitios de unión, uno para proteínas y el otro para la droga anti-cancerosa taxol.
Diagrama de cintas basado en un modelo de una resolución de 3.7 Angstrom desarrollado por científicos del laboratorio de Berkeley, que muestra que la tubulina es un heterodímero que consiste de dos monómeros de estructuras casi idénticas (alfa y beta). Cada monómero está formado por un núcleo de dos láminas beta (verde), rodeado por hélices (azul), y cada uno se une a un nucleótido guanina (rosa). Además de un sitio de unión a un nucleótido, cada monómero tiene otros dos sitios de unión, uno para proteínas y el otro para la droga anti-cancerosa taxol.
El nombre tubulina se refiere una familia de proteínas globulares de 55 Kdalton. La familia de las tubulinas está formada por las tubulinas alfa (α), beta (β) y gamma (γ), que comparten una identidad entre sus cadenas de aminoácidos de 35-40%, aunque su similitud con cualquier otra proteína conocida es mínima.[1] Las tubulinas α y β son las subunidades esenciales de los microtúbulos, mientras que la tubulina-γ es un componente fundamental del centrosoma. Existen asimismo otras variantes menores, que no están presentes en todos los organismos eucariotas, denominadas tubulina-delta (δ), -epsilon (ε) y -zeta (ζ).[2]
Comúnmente, se denomina tubulina a un heterodímero formado por dos subunidades (α y β) que al ensamblarse de manera altamente organizada genera uno de los principales componentes del citoesqueleto, los microtúbulos. Todas las células eucariotas presentan microtúbulos, lo cual indica que las subunidades que los conforman probablemente se originaron cuando los eucariotas aparecieron por vez primera, hace aproximadamente 1.5 billones de años. Es probable que otras proteínas que se asocian a los microtúbulos también daten del origen de los eucariotas, como algunos miembros de las familias de las kinesinas y dineínas, aunque otras son de origen más reciente, como la proteína tau específica de las neuronas.
Contenido
Conservación de las tubulinas α y β
Puesto que las proteínas de la familia de la tubulina tienen un origen muy antiguo, podría esperarse que sus secuencias presentaran una gran divergencia. Sin embargo, esto sólo es cierto para el extremo C-terminal de las tubulinas α y β. Los fragmentos N-terminales están notablemente conservados con variaciones mínimas. Este alto grado de conservación está seguramente impuesto por las limitaciones estructurales del ensamblaje y desensamblaje de microtúbulos (MTs), conjuntamente con las limitaciones impuestas por la asociación de proteínas como las kinesinas y dineínas.[3] Los miembros individuales de la familia de las tubulinas de los distintos órdenes filogenéticos han evolucionado de una manera excepcionalmente lenta, a una tasa comparable la de las histonas o la actina.[4] La alta tasa de conservación dentro de la familia de las tubulinas implica que las propiedades funcionales de estas proteínas imponen unas limitaciones enormes a cualquier diversificación de la secuencia, de manera que las mutaciones sólo pueden acomodarse en unas pocas posiciones sin producir un efecto deletéreo. Por otro lado, una modificación que se haya conservado probablemente sea funcionalmente ventajosa, y por ello estará seguramente relacionada con propiedades específicas de las tubulinas en órdenes diferentes.
Tubulina y estructura de los MTs
Véase también: MicrotúbuloLas interacciones proteína-proteína entre las subunidades de los microtúbulos constituyen una constricción a la estructura terciaria de las tubulinas α y β. Estudios tradicionales de microscopía electrónica utilizando células fijadas con glutaraldehído establecieron que un MT está compuesto normalmente de 13 protofilamentos de tubulina alineados linealmente.[5] Aunque se han identificado variantes de esta estructura, el número de protofilamentos in vivo parece variar entre 12 y 15.[6] El protofilamento consiste de unidades globulares alternadas, con unas dimensiones aproximadas de 50 x 50 x 40 Angstrom, con una unidad que se repite a lo largo del eje longitudinal del protofilamento de alrededor de 80 Angstrom. Estas dimensiones son compatibles con un monómero de tubulina de 50 kD que forma heterodímeros α/β, que constituyen la unidad de 80 Angstrom. Dado que los protofilamentos forman la base del ensamblaje de los MTs, es probable que las interacciones longitudinales entre los heterodímeros sean más estables que las interacciones laterales, entre protofilamentos.
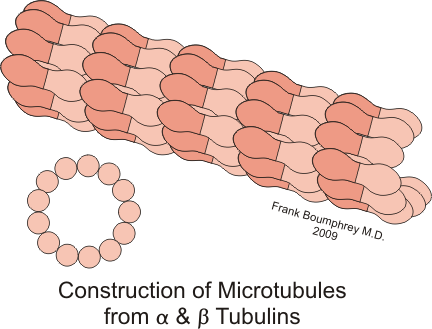
 Esquema de la polimerización de las subunidades de tubulina para formar los microtúbulos.
Esquema de la polimerización de las subunidades de tubulina para formar los microtúbulos.Los heterodímeros se ensamblan en los protofilamentos de tal manera que la tubulina-β de un dímero contacta con la tubulina-α del dímero siguiente. Por ello, los MTs son polares de forma inherente, ya que presentan tubulina-α en un extremo del polímero (el extremo "menos") y tubulina-β en el otro (el extremo "mas"). Los 13 protofilamentos que forman un MT se disponen uno al lado de otro, de manera que si uno sigue las subunidades α o β lateralmente alrededor del MT, se observa que forman una hélice de tres subunidades. Esto significa que la hélice recorre 3 subunidades hasta completar una vuelta. Este tipo de hélice no es perfectamente simétrica, lo que resulta en un una "junta" en la pared del MT en el lugar donde cada hélice completa una vuelta. Los protofilamentos interaccionan entre ellos lateralmente sobre todo a través de contactos α–α y β–β, aunque en la junta la tubulina-α contacta con la tubulina-β.[7] [8]
El dímero de tubulina une 2 moles/mol del nucleótido guanosina: uno está en un sitio intercambiable, mientras que el segundo es no intercambiable. La tubulina purificada de neuronas contiene 1 mol/mol tanto de GTP como de GDP, siendo el GDP el que está unido al sitio intercambiable. El sitio intercambiable está localizado en la subunidad β, y parece que el GTP no intercambiable está unido a la subunidad α.[9] El ensamblaje de MTs es, en casi todas las condiciones, dependiente de GTP o un análogo no hidrolizable de GTP, unido al sitio intercambiable. Esta molécula de GTP se hidroliza posteriormente a GDP, que permanece unido al MT ensamblado y sólo es intercambiable cuando el MT se desensambla. Este cambio en la capacidad de intercambio puede deberse al empaquetamiento de las subunidades en el MT ensamblado o a un cambio de conformación dependiente del ensamblaje.[10]
In vitro, los MTs polimerizan espontáneamente a partir de altas concentraciones de tubulina α y β, en presencia de GTP y Mg2+. El proceso de polimerización ocurre en dos pasos: la "nucleación", que es la etapa limitante, seguida de una rápida elongación.[11] Se piensa que el paso de nucleación implica la formación de un par de protofilamentos cortos, que consistirían de 7, 12 o 18 dímeros de tubulina α/β. Una vez que se ha formado este núcleo, crece rápidamente de manera lateral y longitudinal como una lámina, hasta que se han ensamblado alrededor de 1000 dímeros; en ese momento la lámina se cierra sobre sí misma para formar un cilindro. Las láminas también pueden observarse en los extremos crecientes de MTs preformados, lo que sugiere que los MTs son polímeros de dos dimensiones, y no un polímero helicoidal, en su modo de elongación.[12] Se supone que los MTs se ensamblan de la misma forma in vivo, aunque la concentración de tubulina-α/β dentro de las células está por debajo del nivel necesario para se produzca la nucleación espontánea que se observa in vitro, por lo que el proceso está catalizado por centros organizadores de MTs (COMTs, en inglés MTOCs), como son los centrosomas en las células animales y el cuerpo polar del huso en levaduras. La necesidad de MTOCs in vivo permite a la célula controlar cuándo y dónde se produce la nucleación de MTs. Un gran número de evidencias (experimentos genéticos, estudios de inhibición con anticuerpos, ensayos de complementación in vitro y microscopía de fluorescencia y electrónica) implica la tubulina-γ como la proteína clave responsable de la nucleación de MTs in vivo.[13]
Tubulina-γ
Esta proteína está muy conservada, y es aproximadamente 30% idéntica a las tubulinas α y β, pero no se ensambla en la estructura polimérica de los MTs. Aunque su actividad se concentra en los MTOCs, la mayor parte de la tubulina-γ se encuentra en el citosol. La tubulina-γ citosólica se encuentra principalmente en dos complejos:[14] el complejo mayor en anillo de tubulina-γ (denominado γTuRC, las siglas en inglés de γ-tubulin ring complex) y el complejo pequeño de tubulina-γ (denominado γTuSC, por γ-tubulin small complex), que es una subunidad del γTuRC y es análogo al complejo Tub4 de Saccharomyces cerevisiae.
El complejo γTuRC fue inicialmente aislado a partir de huevos de Xenopus y de células de Drosophila. Consiste aproximadamente de 10–14 moléculas de tubulina-γ y al menos seis proteínas adicionales, generando un complejo de alrededor de 2 MDa. Este complejo está bien conservado, ya que se han identificado complejos similares en células de mamífero. A partir de imágenes de microscopía electrónica, se ha observado que el complejo γTuRC tiene una estructura en anillo abierto y flexible, de alrededor de 25 nm de diámetro.[15] De acuerdo con su estructura, parece ser que este complejo funciona como un molde, a partir del cual crecen los MTs. Como los MTs celulares normalmente contienen 13 protofilamentos, el modelo propone que el γTuRC contiene 13 unidades de tubulina-γ que interaccionan lateralmente. Sin embargo, datos procedentes de varios estudios sugieren que el complejo γTuRC se ensambla a partir de complejos γTuSCs preformados, que contienen dos copias de tubulina-γ y una copia de cada una de las proteínas homólogas de S. cerevisiae Spc97 y Spc98. Esto implica que el complejo γTuRC debe contener un número par de tubulina-γ, en lugar de 13 como se había propuesto inicialmente. Se ha propuesto que los componentes del γTuRC que no forman parte de los γTuSC, forman una "gorra" (cap) que cubre el extremo "menos" de los MTs, que podría regular su actividad, conferir estabilidad a la hélice y/o anclar el γTuRC al centrosoma.
Referencias
- ↑ Little, M.; Seehaus, T. (1988), «Comparative analysis of tubulin sequences», Comp Biochem Physiol B. 90 (4): 655–670, PMID 3073909
- ↑ Oakley, B.R. (2000), «An abundance of tubulins», Trends Cell Biol. 10 (12): 537–542, PMID 11121746
- ↑ Burns, R.G. (1991), «Alpha-, beta-, and gamma-tubulins: sequence comparisons and structural constraints», Cell Motil Cytoskeleton. 20 (3): 181–189, PMID 1773446
- ↑ Doolittle, R.F. (1992), «Reconstructing History With Amino Acid Sequences», Protein Sci 1 (2): 191–200, PMID 1339026
- ↑ Lg, Tilney; Bryan, J.; Bush, D.J.; Fujiwara, K.; Mooseker, M.S.; Murphy, D.B.; Snyder, D.H. (1973), «Microtubules: evidence for 13 protofilaments», J Cell Biol 59 (2): 267–275, PMID 4805001
- ↑ Mogensen, M.M.; Tucker, J.B.; Stebbings, H. (1989), «Microtubule polarities indicate that nucleation and capture of microtubules occurs at cell surfaces in Drosophila», J Cell Biol 108 (4): 1445–52, PMID 2925791
- ↑ Nogales, E.; Whittaker, M.; Milligan, R.A.; Downing, K.H. (1999), «High-resolution model of the microtubule», Cell 96: 79–88, PMID 9989499
- ↑ Nogales, E. (2001), «Structural insights into microtubule function», Annual Review of Biophysics and Biomolecular Structure 30: 397–420, doi:
- ↑ Linse, K.; Mandelkow, E.M. (1988), «The GTP-binding peptide of beta-tubulin. Localization by direct photoaffinity labeling and comparison with nucleotide-binding proteins», J Biol Chem. 263 (29): 15205–10, PMID 3170578
- ↑ Kirchner, K.; Mandelkow, E.M. (1985), «Tubulin domains responsible for assembly of dimers and protofilaments», Embo J. 4 (9): 2397–402, PMID 4076170
- ↑ Voter, W.A.; Erickson, H.P. (1984), «The kinetics of microtubule assembly. Evidence for a two-stage nucleation mechanism», J. Biol Chem. 259: 10430–10438, http://www.jbc.org/cgi/content/abstract/259/16/10430, consultado el 2009-07-28
- ↑ Chretien, D.; Fuller, S.D.; Karsenti, E. (1995), «Structure of growing microtubule ends: two-dimensional sheets close into tubes at variable rates», J Cell Biol 129: 1311–1328, http://jcb.rupress.org/cgi/content/abstract/129/5/1311, consultado el 2009-07-28
- ↑ Moritz, M.; Agard, D.A. (2001), «γ-Tubulin complexes and microtubule nucleation», Current Opinion in Structural Biology 11: 174–181, doi:
- ↑ Schiebel, E. (2000), «γ-Tubulin complexes: binding to the centrosome, regulation and microtubule nucleation», Curr Opin Cell Biol 12: 113–118, doi:
- ↑ Oegema, K.; Wiese, C.; Martin, O.C.; Milligan, R.A.; Iwamatsu, A.; Mitchison, T.J.; Zheng, Y. (1999), «Characterization of two related Drosophila γ-tubulin complexes that differ in their ability to nucleate microtubules», J Cell Biol 144 (721): 721–733, http://jcb.rupress.org/cgi/content/abstract/144/4/721, consultado el 2009-07-28
Categoría:- Proteínas
Wikimedia foundation. 2010.