- Lanzadera de citrato
-
La lanzadera de citrato-malato, o citrato-piruvato, es un mecanismo de transporte de ácido acético que encontramos en la membrana mitocondrial interna, y que permite que esta molécula pueda ser utilizada en el citosol celular. Este acetilo se forma en importantes vías catabólicas como la beta oxidación de ácidos grasos o en la descarboxilación oxidativa del piruvato, reacción clave catalizada por el complejo piruvato deshidrogenasa que actúa de puente entre la glicólisis y el ciclo de Krebs, ambas tienen lugar en el interior de la mitocondria. El acetil-CoA es también una molécula clave en diversas vías anabólicas como lo síntesis de ácidos grasos, aminoácidos, acetilcolina o en la gluconeogénsis, reacciones que en su gran mayoría tienen lugar en el citosol, y por lo tanto requieren la presencia de esta molécula fuera de las mitocondrias.
Tiene ímpetu la cosa
Contenido
Conceptos básicos
Acetil-CoA es la forma habitual en la que el ácido acético está presente en el cuerpo. La membrana mitocondrial interna es impermeable al acetil-CoA, por lo tanto ésta molécula no puede ser transportada a través de la mitocondria mediante difusión simple. Además, el acetato, a diferencia de los ácidos grasos, no puede ser transportado a través de carnitina. Por todas esta razones el acetato necesita un mecanismo de transporte independiente del resto, es decir, exclusivo para esta molécula clave en el metabolismo energético.
Mecanismo
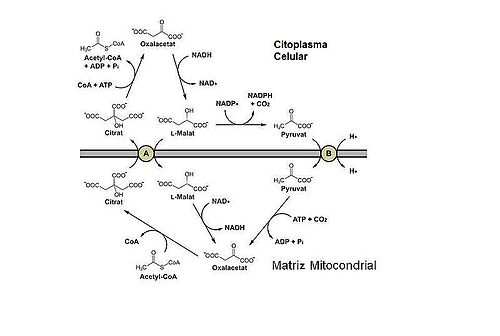
El acetil-CoA intramitocondrial reacciona primeramente con oxalacetato para formar citrato, una reacción catalizada por la citrato sintasa, que corresponde a la primera reacción del ciclo de Kreps.
Acetil-CoA + oxalacetato -------> Citrato + CoASH
La enzima citrato sintasa fue descrita por Severo Ochoa quien la denominó como enzima condensante, pues lleva a cabo una condensación aldólica entre el metilo del Acetil-CoA y el carbonilo del oxaloacetato. La citrato sintasa es inhibida por succinil-CoA, ATP, NADPH, ésteres de CoA y ácidos grasos de cadena larga (18C), no se sabe si esto último tiene significado biológico. Cabe destacar que se trata de una reacción de gran importancia en el ciclo de Kreps, y sucede que cuándo esta vía central del catabolismo energético es inhibida, principalmente cuándo la concentración de ATP es elevada o suficiente, el exceso de citrato en la matriz se utiliza para transportar acetil-CoA al citoplasma, donde puede ser utilizado para la síntesis de ácidos grasos y colesterol. Adicionalmente, los niveles altos de citrato en el citoplasma activan la enzima regulatoria clave de la síntesis de los ácidos grasos, acetil-CoA carboxilasa. En tejidos no hepáticos el citrato se requiere también para la síntesis de cuerpos cetónicos.
Entonces el citrato generado es trasportado al citosol mediante la lanzadera de citrato, proceso en el que se introduce malato en el interior de la mitocondria. Esta lanzadera es un antiporte, es decir, una proteína integral, de membrana, que transporta dos moléculas (en este caso malato y citrato) en direcciones opuestas. Existen gran cantidad de proteínas de membrana con esta función, en la membrana mitocondrial destacan la lanzadera de malato-aspartato, la lanzadera de glicerolfosfato o el transportador de piruvato, entre otras.
En el citosol, el citrato regenera acetil-CoA en una reacción dependiente de ATP. Esta reacción es la inversa a la producida en la matriz mitocondrial, por ello también se genera oxaloacetato.
Citrato + CoA + ATP -------> acetil-CoA + oxaloacetato + ADP + Pi
La enzima que cataliza esta reacción es la ATP-citrato liasa. La reacción inversa es una reacción exergónicas, ya que la ruptura del enlace energético del CoA, permite la liberación de gran cantidad de energía. En este caso, proceso inverso, éste enlace energético ha de formarse, por lo que se requiere la intervención del ATP, que gracias a la ruptura de uno de sus dos enlaces esterfosfóricos, permite la regeneración del Acetil-CoA. Por lo tanto, el acetil-CoA ya se encuentra disponible en el citosol para participar en distintas vías anabólicas.
El oxaloacetato generado en esta reacción no puede ser devuelto directamente al interior de la mitocondria ya que la membrana mitocondrial carece de un transportador específico para esta molécula. Por esta razón el oxalacetato generado se reduce a malato, gracias a la malato deshidrogenasa, de tal manera que puede retornar a la matriz mitocondrial gracias a la lanzadera de citrato.
oxalacetato + NADH + H+ -------> malato + NAD+
Para completar el ciclo de la lanzadera de citrato-malato, el malato una vez introducido en la matriz es oxidado a oxalacetato, permitiendo de este modo que puede reiniciarse el proceso. Esta reacción es catalizada por la enzima malato deshidrogenasa y destaca por la reducción del NADH, recuperando de éste modo el NADH que se había requerido en el citosol en el proceso de reducción del oxalacetato a malato.
Hay que destacar que el malato producido en el citosol a partir de la reducción del oxalacetato puede invertirse para la generación de NADPH mediante una reacción de descarboxilación, proceso en el que se libera dióxido de carbono y piruvato. Ésta reacción es catalizada por una enzima malato deshidrogenasa específica unida a NADP, que no tan solo oxida el malato, proceso que originaria únicamente oxalacetato i NADH, sino que también lo descarboxila, dando lugar al piruvato y el dióxido de carbono. Esta reacción es una importante fuente de NADPH en el tejido adiposo y el hígado, cuando se sintetizan ácidos grasos a partir de glucosa.
Por lo tanto, el NADPH no se obtiene tan solo en la vía de las pentosas fosfato sino que también se produce en este paso de malato a piruvato en el citosol, siendo estas dos, las únicas formas de obtenerlo.
El piruvato formado en el citosol a partir de malato puede introducirse posteriormente en la matriz mitocondrial gracias al antiporte, que extrae protones de la mitocondria a costa de introducir piruvato en ella. El piruvato es capaz de generar oxalacetato mediante una reacción dependiente de ATP.
Piruvato + CO2 + H2O + ATP -------> Oxalacetato + ADP + Pi + 2H+
Vemos que el producto de la glicólisis, el piruvato, también sirve como proveedor de importantes componentes biosintéticos. Esta reacción es catalizada por la piruvato carboxilasa, una enzima activada por el acetil-CoA, que indica la falta de oxalacetato. Se trata de una reacción anaplerótica.
Por lo tanto, cuando se completa un ciclo citrato piruvato se desplaza una acetil-CoA al citosol, se obtiene NADH en el interior de la mitocondria y NADPH en el exterior, pero para este proceso es necesario consumir dos moléculas de ATP, una en la carboxilación del piruvato, y otra en la ruptura del citrato. Aún así este ciclo no siempre se realiza de este modo, es decir, no siempre se forma piruvato a partir del citrato DIANA PUTÓN, de tal manera que el ciclo finalizaría con la producción de malato en el citosol, que seria introducido por la lanzadera a la matriz mitocondrial, y entonces tan solo seria necesaria una molécula de ATP por cada ciclo.
Enlaces externos
http://www.ciclodekrebs.com/etapas_del_ciclo_de_krebs
http://www.monografias.com/trabajos46/acido-citrico-krebs/acido-citrico-krebs2.shtml
Bibliografía
- MULLER ESTERL, WERNER · REVERTE; 2008; 01 ed. ISBN: 978-84-291-7393-2
- LEHNINGER PRINCIPIOS DE BIOQUiMICA Editorial Omega 8428214107 9788428214100.


Wikimedia foundation. 2010.