- Physeter macrocephalus
-
 Cachalote
Cachalote
Cachalote
Estado de conservación 
Vulnerable (UICN)[1]Clasificación científica Reino: Animalia Filo: Chordata Clase: Mammalia Orden: Cetacea Suborden: Odontoceti Familia: Physeteridae Género: Physeter Especie: P. macrocephalus Nombre binomial Physeter macrocephalus
Linnaeus, 1758Distribución 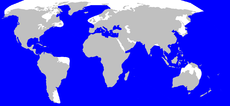
El cachalote (Physeter macrocephalus) es una especie de mamífero marino del orden Cetácea del suborden Odontoceti, que posee el cerebro más grande que cualquier otro animal existente. El cachalote es el único miembro del género Physeter. Es una de las tres especies vivientes de la familia Physeteridae, junto con el cachalote pigmeo y cachalote enano.
Un macho puede crecer hasta 20,5 metros de largo. Es el animal dentado más grande que existe. La cabeza puede tener hasta un tercio de la longitud total del animal. Tiene una distribución cosmopolita en todos los océanos. La especie se alimenta de calamares y peces, sumergiéndose a profundidades de hasta tres kilómetros, lo cual lo convierte en el mamífero que se zambulle a mayor profundidad. Su dieta incluye el calamar gigante y el calamar colosal. Es el depredador viviente más grande y posiblemente el más grande que jamás haya existido, no en términos de comer animales (lo cual es cierto en todos los cetáceos, incluyendo las grandes ballenas) sino en que depreda activamente animales autónomos. El chasquido producido por el cachalote es el sonido más intenso producido por animal alguno, pero su función es desconocida. Esta ballena vive en grupos, en los cuales las hembras y sus crías viven separados de los machos más viejos. Las hembras cooperan para proteger y alimentar sus retoños. Dan a luz cada tres a seis años, y el cuidado de las crías lleva más de una década.
El término cachalote deriva de la palabra francesa antigua usada para «diente». Durante el periodo comprendido entre principios del siglo XVII hasta finales del siglo XX, el cachalote fue cazado para obtener el espermaceti y otros productos, como aceite y ámbar gris. Al espermaceti se le encontró un uso amplio, como insumo para velas, jabón, cosméticos y aceite para uso industrial. Debido a su tamaño, podía algunas veces defenderse eficazmente de los primeros balleneros. El caso más famoso fue el barco Essex hundido por un cachalote en 1820. Como resultado de la caza intensiva en los siglos XIX y XX se encuentra actualmente señalado como especie vulnerable por la UICN. El cachalote tiene pocos depredadores naturales, ya que pocos animales son lo suficientemente fuertes para atacar con éxito a un adulto saludable, sin embargo las orcas atacan los grupos y matan sus crías. Son animales longevos y pueden vivir más de 70 años.
Contenido
Nombre
En inglés el cachalote es llamado sperm whale un apócope de spermaceti whale (ballena de espermaceti). En español el nombre «cachalote» se piensa que deriva del francés arcaico «diente», similar a «cachau» en el dialecto gascón.[2] El diccionario etimológico de Corominas dice que el origen de la palabra es incierto, pero se sugiere que viene del latín vulgar «cappula», plural de «cappulum», empuñadura de espada.[3] De acuerdo al diccionario Encarta, la palabra «cachalot» vino al inglés «vía francés del español o tal vez del portugués, cachola que significa cabeza grande».
Descripción
Tamaño
Tamaño promedio [4] Longitud Peso Macho 16 m 41.000 kg Hembra 11 m 14.000 kg Recién nacido 4 m 1.000 kg El cachalote es la ballena dentada más grande, con machos adultos de 20,5 metros de longitud máxima y peso de 57.000 kilogramos.[5] [6] La especie de ballena dentada que le sigue en tamaño es el zifio de Baird, con 12,8 metros de largo y 15.000 kilogramos de peso.[7] El museo de ballenas en la isla de Nantucket (Massachusetts, Estados Unidos de Norteamérica) tiene una mandíbula de 5,5 metros de longitud. En el museo se afirma que este individuo tenía 24 metros de largo; mientras que el cachalote que hundió el ballenero Essex (uno de los incidentes que inspiró Moby-Dick) se estimó que tenía 26 metros.[8] [9] La cacería intensiva pudo haber disminuido el tamaño promedio de los cachalotes, puesto que los machos grandes eran los más buscados y capturados, principalmente después de la Segunda Guerra Mundial.[9] En la actualidad, los machos no exceden con frecuencia los 18,3 metros de longitud y 51.000 kilogramos de peso.[4]
Se encuentra entre los cetáceos que presentan un dimorfismo sexual más evidente. Al momento del nacimiento ambos sexos tienen casi el mismo tamaño,[4] pero los machos maduros tienen usualmente entre 30 a 50 por ciento mayor longitud y tres veces más peso que las hembras.[5]
Apariencia
El aspecto distintivo del cachalote proviene de su enorme cabeza, la cual frecuentemente tiene un tercio de la longitud del animal. El espiráculo se localiza muy próximo al extremo frontal de la cabeza, desplazado a la izquierda de la línea media.[5]
 Aletas posteriores de un cachalote al sumergirse en el golfo de México.
Aletas posteriores de un cachalote al sumergirse en el golfo de México.Las aletas posteriores del cachalote son triangulares y muy gruesas. Las levanta siempre fuera del agua cuando inicia una inmersión.[5] Tiene una serie de crestas en el tercio caudal en lugar de una aleta dorsal. La cresta más larga fue llamada «hump» (joroba) por los balleneros, y puede ser confundida con una aleta dorsal debido a su forma.[4]
Al contrario de la piel lisa de la mayoría de las grandes ballenas, la piel de la espalda es con mucha frecuencia nudosa y ha sido comparada con una ciruela por los observadores entusiastas.[10] La piel tiene normalmente un color gris uniforme, sin embargo puede parecer marrón bajo la luz del sol. También se han reportado ejemplares albinos.[11] [12] [13]
Mandíbulas y dientes
El cachalote tiene 20 a 26 dientes en cada lado de la mandíbula inferior.[5] Los dientes tienen forma cónica y pesan hasta un kilogramo.[14] El propósito de los dientes es desconocido, pues no parecen ser necesarios para capturar o comer calamares, ya que se han observado animales bien alimentados que no tenían dentadura. Una hipótesis es que los dientes son usados para la agresión entre machos.[15] Los machos a menudo muestran cicatrices que parecen ser causadas por dientes. En la mandíbula superior cuentan con dientes rudimentarios, pero raramente emergen a la cavidad oral.[16]
Respiración e inmersión
Los cachalotes, junto con los zifios calderones y los elefantes marinos, son los mamíferos que se sumergen más profundo.[5] Se cree que son capaces de alcanzar los tres kilómetros de profundidad y permanecer sumergidos 90 minutos.[5] [17] Con mayor frecuencia bucea a una distancia de 400 metros de la superficie durante 35 minutos.[5] A esta gran profundidad en ocasiones se han enredado y ahogado con cables submarinos.[18]
 Cachalote arqueando la espalda preparándose para sumergirse en Dominica.
Cachalote arqueando la espalda preparándose para sumergirse en Dominica.
El cachalote se ha adaptado para arreglárselas con los cambios drásticos de presión cuando se sumerge. Su caja torácica flexible permite el colapso pulmonar, reduciendo la entrada de nitrógeno, y el metabolismo puede disminuir para conservar oxígeno.[19] [20] La mioglobina, una proteína que almacena oxígeno en el músculo, es mucho más abundante que en los animales terrestres.[21] La sangre tiene alta densidad de glóbulos rojos, los cuales contienen el transportador de oxígeno hemoglobina. La sangre oxigenada puede dirigirse hacia el cerebro y a otros órganos esenciales solamente cuando los niveles de oxígeno disminuyen.[22] [23] [24] El órgano del espermaceti también puede jugar un papel para ajustar la flotabilidad.[25]
A pesar que los cachalotes se encuentran bien adaptados para el buceo de profundidad, las inmersiones repetidas tienen efectos adversos a largo plazo. Los huesos muestran lesiones causadas por la descompresión rápida. Los esqueletos de individuos viejos muestran los daños más extensos, mientras los esqueletos de individuos jóvenes no evidencian ninguna lesión. Estos daños pueden indicar que estas ballenas son susceptibles a la descompresión y el ascenso súbito a la superficie puede ser letal para ellas.[26]
Entre inmersiones, sube a respirar por unos 8 minutos antes de la siguiente inmersión.[5] Como los demás odontocetos respiran a través de un único espiráculo en forma de «S». Respiran de 3 a 5 veces por minuto en reposo, incrementándose hasta 6 a 7 veces por minuto después de una inmersión. El soplido es único y ruidoso, con un chorro que se eleva hasta 15 metros sobre la superficie y apunta hacia delante y la izquierda en un ángulo de 45°. En promedio, las hembras y jóvenes respiran cada 12,5 segundos antes de sumergirse, mientras que los machos grandes lo hacen cada 17,5 segundos antes de la inmersión.[27]
Cerebro y sentidos
El cerebro del cachalote es el más grande de cualquier animal moderno o extinto, pesando un promedio de 8 kilogramos.[28] [29] Sin embargo no es muy grande en proporción al tamaño del cuerpo. Por ejemplo, los cachalotes tienen un cociente de encefalización menor que muchos delfines y ballenas, más bajo que los grandes simios y mucho menor que los humanos.[29] [30]
Como otros odontocetos, los cachalotes usan la ecolocalización como un medio para encontrar comida debido a que su hábitat tiene condiciones acústicas favorables y la absorción por el agua y turbidez limita el rango visual. La ballena emite chasquidos de alta frecuencia enfocados en un ángulo amplio. Genera sonidos pasando aire desde los orificios nasales óseos a través de los labios fónicos, una estructura dentro de la cabeza.[31] El cráneo, el melón y varios sacos de aire en la cabeza de la ballena, juegan un papel importante en la transmisión de los sonidos y la mandíbula inferior como una vía para la recepción de los ecos. Un canal continuo lleno de grasa trasmite los sonidos al oído interno.[32]
Función del espermaceti
El órgano del espermaceti puede ayudar a ajustar la flotabilidad de las ballenas. Antes de la inmersión, el agua fría hace contacto con la cavidad y solidifica la cera.[25] [33] El incremento en la densidad específica genera una fuerza descendente de unos 40 kilogramos y permite a la ballena descender con menos esfuerzo. Durante la cacería, el consumo de oxigeno produce calor y derrite el espermaceti, incrementando la flotabilidad, facilitándole el regresos a la superficie.[34]
Herman Melville en su novela Moby-Dick sugiere que el órgano del espermaceti evolucionó como un tipo de ariete para embestir durante las luchas entre machos.[35] Esta hipótesis es consistente con los hundimientos, bien documentados, de los barcos balleneros Essex y Ann Alexander por atacantes con un peso estimado, de máximo, una quinta parte del peso de los navíos.[36]
Otra posibilidad es que el órgano ayude a la ecolocalización. La variación en la forma del órgano reduce o amplifica el sonido.[37]
El cachalote tiene dos orificios nasales. Una fosa externa forma el espiráculo, y una interna presiona contra el compartimiento que contiene el espermaceti. El órgano del espermaceti del macho es mucho más grande que el de las hembras. Esto puede ser un caso de selección sexual, impulsando los machos a competir por las hembras usando despliegues sonoros.[38]
Ecología, comportamiento y modo de vida
Distribución
El cachalote se encuentra entre las especies más cosmopolitas. Prefiriendo las aguas no congeladas hasta los 1000 metros de profundidad.[1] Sin embargo ambos sexos se movilizan a través de océanos y mares tropicales y templados, solo los machos adultos pueblan las latitudes más altas.[11]
Abunda relativamente desde los polos al ecuador, y se encuentra en todos los océanos. Habita el mar Mediterráneo pero no el mar Negro,[4] mientras su presencia en el mar Rojo es incierta.[1] Las entradas de baja profundidad tanto del mar Negro como el mar Rojo puede contribuir a su ausencia.[39] Las capas inferiores del mar Negro carecen de oxígeno y contienen altas concentraciones de compuesto de sulfuros como el sulfuro de hidrógeno.[40]
Las poblaciones son densas cerca a las plataformas continentales y cañones submarinos.[11] Los cachalotes son encontrados usualmente en aguas profundas de alta mar, pero pueden hallarse cerca a las costas en áreas donde la plataforma continental es pequeña y baja rápidamente a profundidades entre 310 y 920 metros.[4] Las áreas costeras con poblaciones considerables de estas ballenas son las islas Azores y la isla caribeña de Dominica.[41]
Reproducción
 Cachalote joven.
Cachalote joven.Los cachalotes pueden vivir 70 años o incluso más.[4] [11] [42] Son el ejemplo ideal de una especie que ha sido sometida a selección K, una estrategia reproductiva asociada con condiciones ambientales estables, una baja tasa de natalidad, ayuda significativa de los parientes hacia las crías, maduración lenta y alta longevidad.[5]
No se ha determinado todavía como eligen sus parejas. Hay evidencia de que los machos tienen jerarquías y existe también evidencia que esto influye en la selección para el apareamiento.[43] La gestación requiere 14 a 16 meses, produciendo solo una cría.[4] La lactancia trascurre hasta los 19 a 42 meses, pero los jóvenes pueden succionar hasta los 13 años.[4] Las crías pueden alimentarse de hembras diferentes a su madre.[4] Las hembras generalmente tienen intervalos entre nacimientos de tres a seis años.[4]
Las hembras alcanzan la madurez sexual entre los 7 a los 13 años, y los machos la alcanzan a los 18 años. Una vez alcanzada la madurez, los machos se trasladan a latitudes más altas, donde el agua es más fría y la alimentación es mejor. Las hembras permanecen en latitudes bajas.[4]
Los machos alcanzan el mayor tamaño hacia los 50 años.[5]
Comportamiento social
 Diagrama de la formación en margarita.
Diagrama de la formación en margarita.Las hembras permanecen en grupos de aproximadamente una docena de individuos con sus crías.[5] Los machos integran estas «guarderías» en forma permanente entre los 4 a 21 años de edad y forman un «grupo de solteros» con otros machos de edad y tamaño similares.[5] Cuando los machos maduran, tienden a dispersarse en grupos más pequeños y los más viejos en su mayoría tienen vidas solitarias.[5] Machos maduros se han varado juntos, lo cual sugiere un grado de cooperación, que aun no ha sido totalmente entendido.[5]
El atacante no humano más común de los cachalotes es la orca, pero las ballenas piloto y las falsas orcas también algunas veces los atacan o acosan.[44] [45] Las orcas amenazan los grupos de hembras con crías, usualmente intentando aislar y matar un individuo joven. Las ballenas hembras repelen estos ataques rodeando sus vástagos. Los adultos usan sus aletas de cola contra las orcas dando la cara a las crías, o de frente, luchando con sus dientes.[5] Esta formación en margarita es también empleada para auxiliar un miembro lesionado de la manada, un comportamiento que los primeros balleneros explotaron, atrayendo las demás ballenas hiriendo solo uno de los miembros del grupo.[46] Si el grupo de orcas es muy grande, estas pueden algunas veces matar hembras adultas. Los ejemplares más grandes no tienen predadores aparte de los humanos, y se cree que son demasiado grandes y fuertes para ser amenazados por las orcas.[47]
Alimentación
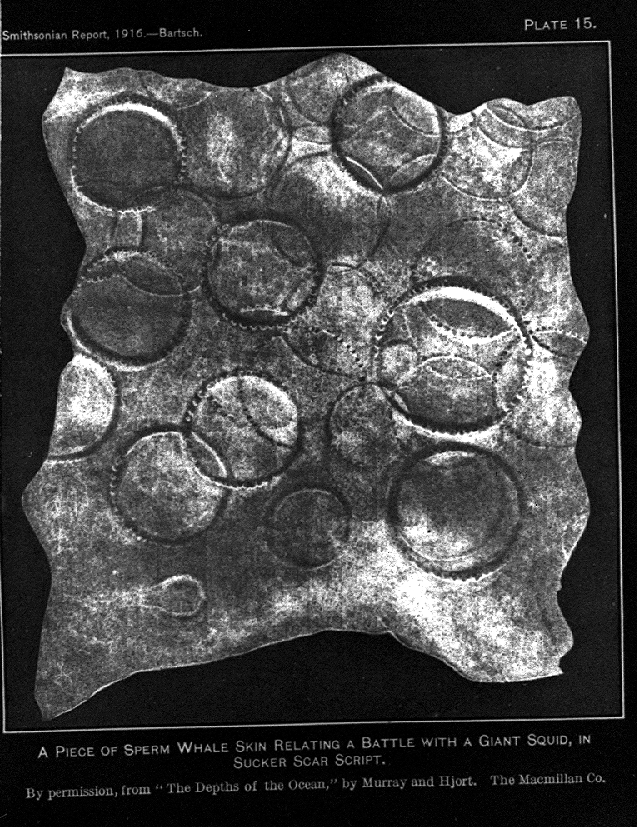
 Un fragmento de piel de cachalote con cicatrices de ventosas de calamar gigante.
Un fragmento de piel de cachalote con cicatrices de ventosas de calamar gigante. Ataque a su presa.
Ataque a su presa.Los cachalotes habitualmente bucean entre los 300 a 800 metros de profundidad, y algunas veces hasta 1 a 2 kilómetros para buscar comida.[48] Tales inmersiones pueden durar más de una hora.[48] Se alimentan de varias especies, de las cuales las más notables son el calamar gigante, el calamar colosal, los pulpos, y diversos peces como rayas en la zona demersal, pero la mayor parte de su dieta consiste en calamares de mediano tamaño.[49] Algunas especies pueden ser capturadas incidentalmente mientras la ballena se está alimentando de calamares.[49] La mayor parte de lo que se conoce de los calamares de las profundidades ha sido aprendido de ejemplares en el estómago de cachalotes capturados, como también, en estudios más recientes mediante el análisis de su materia fecal. En un estudio llevado a cabo en inmediaciones de las islas Galápagos, se encontró que los cachalotes habían ingerido con mayor frecuencia calamares de los géneros Histioteuthis (62%), Ancistrocheirus (16%) y Octopoteuthis (7%) pesando cada uno entre 12 a 650 gramos.[50] Las supuestas batallas con los calamares colosales (los cuales pesan 500 kilogramos) nunca han sido observadas por humanos; sin embargo las cicatrices en su cuerpo se cree que son la mayoría causadas por grandes calamares.
Un estudio de 1966 que examinaba estas ballenas, que habían sido capturadas por los balleneros neozelandeses en la región del estrecho de Cook, se encontró una relación 1,69:1 en peso, entre calamares y peces ingeridos.[51] Los cachalotes algunas veces sustraen bacalaos y merluzas de las líneas de pesca. Los pescadores de línea larga en el golfo de Alaska se quejan que los cachalotes toman ventaja de sus operaciones pesqueras para comer especies apetecidas sustrayéndolas de las líneas, eludiendo la necesidad de cazar.[52] Sin embargo la cantidad de pescado tomado es muy pequeño comparado con los requerimientos diarios de este animal. Una filmación de video reciente captó un gran ejemplar macho «rebotando» una línea larga, para obtener pescado.[53] Se cree que los cachalotes depredan la especie de tiburón llamado tiburón de boca ancha, una especie rara y grande de las profundidades, descubierto en 1970.[54] [55]
En una ocasión tres cachalotes fueron observadas atacando o jugando con un boca ancha.[56]
La irritación producida en el intestino por el pico afilado engullido de los calamares, puede iniciar la producción de una sustancia lubricante que da lugar a la formación de ámbar gris, en forma análoga a la producción de las perlas.[57] Los cachalotes son comensales prodigiosos y comen alrededor del 3 por ciento de su peso corporal por día. El consumo anual de presas por estos cetáceos alrededor del mundo se estima en cerca de 100.000.000 de toneladas, una cantidad más alta que el consumo total de animales marinos por los humanos cada año.[58]
No se encuentra claro porque la cabeza del cachalote, es tan grande en comparación con la mandíbula inferior. Una teoría es que la habilidad para la ecolocación se ubica en su cabeza, situación que le ayudaría en la cacería. Sin embargo los calamares, su principal presa, pueden tener propiedades acústicas muy similares al agua de mar para reflejar sonidos.[59] En el interior de la cabeza contiene unas estructuras llamadas labios fónicos, a través del cual se impulsan los flujos de aire para emitir sonidos. Pueden crear un chasquido que tienen una intensidad de 230 decibelios medida a la distancia de 1 metro, en otras palabras, es mucho más fuerte que el sonido hecho por cualquier otro animal, y 10-14 decibelios más fuerte que el sonido de un fusil ubicado en el aire a 1 metro de distancia.[60] Se ha sugerido que los chasquidos sirven para aturdir a sus presas. Los estudios experimentales que tratan de probar este efecto, no han sido capaces de reproducir esta acción sobre sus presas, poniendo en duda esta idea.[61]
Taxonomía y denominación
El cachalote pertenece al orden Cetacea, a este orden pertenecen todas las ballenas y delfines. Es miembro del suborden Odontoceti, el cual engloba todas las ballenas dentadas y delfines. Es la única especie existente del género, Physeter, en la familia Physeteridae. Dos especies existentes del género Kogia, el cachalote pigmeo Kogia breviceps y el cachalote enano K. simus, están ubicados en la familia Physeteridae. Algunas clasificaciones ubican el género Kogia en la familia Kogiidae.[62] En otros esquemas taxonómicos las familias Kogiidae y Physeteridae son combinadas en la superfamilia Physeteroidea.[63]
El cachalote fue una de las especies originalmente descrita por Linneo en 1758 en su obra del siglo XVIII, Systema Naturae. Él reconoció cuatro especies en el género Physeter.[64] Los expertos pronto concluyeron que existía solo una especie, sin embargo había un debate sobre si debía ser llamado P. catodon o P. macrocephalus, dos de los nombres usados por Linneo. Ambos nombres permanecen vigentes, sin embargo los autores más recientes aceptan como nombre valido macrocephalus, limitando el nombre P. catodon a sinonimia.[a]
Historia evolutiva
Registro fósil
A pesar que el registro fósil de los cachalotes es pobre,[65] algunos géneros extintos han sido asignados a la superfamilia Physeteroidea, la cual incluye los últimos ancestros comunes del cachalote moderno, del cachalote enano y cachalote pigmeo. Estos fósiles incluyen los géneros Ferecetotherium, Idiorophus, Diaphorocetus, Aulophyseter, Orycterocetus, Scaldicetus, Placoziphius, Zygophyseter y Acrophyseter.[59] [63] [66] Ferecetotherium, encontrado en Azerbaiyán y fechado en el Oligoceno tardío (hace 28 a 23 millones de años), es el fósil más primitivo que se ha encontrado, con características del cachalote, como el rostro asimétrico («pico» o «trompa»).[67] La mayoría de los fósiles datan del periodo Mioceno, 23 a 5 millones de años. Diaphorocetus, de Argentina, ha sido fechado en el Mioceno Temprano. Los fósiles de cachalote del Mioceno Medio, incluyen los géneros Aulophyseter, Idiorophus y Orycterocetus, todos ellos encontrados en la costa este de los Estados Unidos, y Scaldicetus, encontrado en Europa y Japón.[67] [68] Fósiles del género Orycterocetus se han encontrado también en el océano Atlántico norte y en el mar Mediterráneo, adicionalmente a los encontrados en la costa oeste de los Estados Unidos.[69] Placoziphius, encontrado en Europa, y Acrophyseter, en el Perú, están fechados en el Mioceno Tardío.[63] [67]
El árbol evolutivo familiar de los cachalotes,[70]Cetacea Odontoceti Physeteroidea Otros Physeteroidea†
Kogiidae Cachalote pigmeo
Cachalote enano
Physeteridae Otros Physeteridae†
Cachalote
incluye un resumen simplificado de los grupos extintos (†)[59]Los fósiles encontrados difieren de los modernos, en la fórmula dental y la forma de la cara y mandíbulas.[67] Por ejemplo Scaldicetus tenía un rostro cónico.[68] Los géneros del Oligoceno y el Mioceno Temprano y Medio, con la posible excepción de Aulophyseter, tenían dientes en las mandíbulas superiores.[67] Acrophyseter, del Mioceno Tardío, también tenía dientes tanto en la mandíbula superior como inferior, un rostro corto y una mandíbula inferior con una curva ascendente.[63] Estas diferencias anatómicas sugieren que estas especies fósiles no necesariamente se alimentaban de calamares de las grandes profundidades como el cachalote moderno, en lugar de ello consumirían pescado.[67] Zygophyseter, vivió en el Mioceno Medio a Tardío, encontrado en el sur de Italia, tenía dientes en ambas mandíbulas y parecían estar adaptados a comer grandes presas, como lo hacen las orcas modernas.[59]
Filogenia
La postura tradicional es que los misticetos y los odontocetos provienen de las ballenas primitivas del periodo Oligoceno Temprano, y que la superfamilia Physeteroidea, el cual incluye el cachalote y el cachalote enano y pigmeo, se separaron de las otras ballenas dentadas poco después, hace 23 millones de años.[65] [67] Entre los años 1993 a 1996 mediante análisis de filogenia molecular realizados por Milinkovitch y colegas, basados en la comparación de genes de varias ballenas modernas, sugirieron que los cachalotes, se encuentran más estrechamente relacionadas a los misticetos que a las otras ballenas dentadas, lo cual podría significar que los odontocetos no son monofiléticos, o sea que no todos son descendientes de una sola ballena ancestral dentada.[70] Sin embargo algunos estudios más recientes, basados en varias combinaciones de anatomía comparada y filogenia molecular, han desmentido los análisis de Milinkovitch por motivos técnicos y reafirmado que los odontocetos son monofiléticos.[70] [71] [72]
Estos análisis también confirmaron que se produjo una rápida radiación adaptativa de los physiteroideos en el Mioceno.[59] Los kógidos (cachalote enano y pigmeo) se separaron de Physeteridae hace al menos 8 millones de años.[71]
Relación con humanos
Caza histórica
El espermaceti, en su totalidad obtenido del órgano del espermaceti, y el aceite de cachalote, obtenido de la grasa corporal, eran muy codiciados por los balleneros durante los siglos XVIII, XIX y XX. Esta sustancia tenía una variedad de aplicaciones comerciales, para velas, jabones, cosméticos, aceite industrial y otros lubricantes especiales, aceite para lámparas, lápices, crayones, impermeabilizante para cueros, materiales anticorrosivos y muchos compuestos farmacéuticos.[73] [74] [75] [76] El ámbar gris, una sustancia sólida, serosa e inflamable producida en el sistema digestivo de los cachalotes, era usado también como un fijador en perfumería.
 Cacería de cachalotes.
Cacería de cachalotes.Antes de principios del siglo XVIII, la cacería era realizada principalmente por nativos indonesios.[77] La leyenda cuenta que en algún momento a principios del siglo XVIII alrededor del año 1712, el Capitán Christopher Hussey, mientras buscaba ballenas francas cerca a la costa, fue desviado por el viento hacia el Norte, donde encontró una manada de cachalotes y mato uno.[78] A pesar que la historia puede que no sea real, los balleneros americanos iniciaron muy pronto la caza de estos animales. El juez Paul Dudley, en su Ensayo sobre la historia natural de las ballenas (1725), afirma que un hombre de apellido Atkins, quien duró diez o veinte años en el negocio, estuvo entre los primeros que cazo uno, alrededor de 1720 frente a la costa de Nueva Inglaterra.[79]
Existen solo unos pocos casos registrados de cacería de cachalotes en altamar en el periodo comprendido entre 1709 hasta 1730. Debido a que las balandras apostadas en los bancos de Nantucket, capturaban ballenas francas o iban a la región del estrecho de Davis a cazar ballenas de Groenlandia. A principios de 1740, con el desarrollo de las velas de espermaceti, los buques estadounidenses empezaron a concentrarse en estos cetáceos. El diario de Benjamin Bangs (1721-1769) narra que navegando en su balandra, el encontró otros tres barcos destazando cachalotes de la costa de Carolina del norte a finales de mayo de 1743.[80] De regreso a Nantucket en el verano de 1744 en un viaje posterior el anotó «45 espermacetos fueron traídos aquí este día», otra indicación de que los balleneros norteamericanos estaban en pleno apogeo.[80]
Los balleneros pronto abandonaron las costas de las colonias americanas, hacia la corriente del golfo, los Grandes Bancos de Terranova, África Occidental (1763), las islas Azores (1765) y el Atlántico sur (1770). De 1770 a 1775 los puertos de Massachusetts, New Cork, Connecticut y Rhode Island produjeron 45.000 barriles de aceite de cachalote anualmente, en comparación a los 8.500 de aceite de otras ballenas.[81] En esta misma época los británicos empezaron la cacería de cachalotes, empleando barcos y personal americanos.[82] Para la década siguiente los franceses entraron al negocio, también empleando a expertos americanos.[82] La cacería de estas ballenas se incrementó hasta mediados del siglo XIX. El espermaceti fue importante en el alumbrado público (por ejemplo fue usado para alumbrar viviendas, siendo usado hasta 1862, cuando fue reemplazado por aceite de manteca de cerdo, que a su vez fue reemplazado por petróleo) y para lubricación de maquinaria (como las usadas en fabricas de algodón) de la revolución industrial. La cacería declinó en la segunda mitad del siglo XIX, cuando el petróleo se empezó a utilizar ampliamente. es este sentido, puede decirse que protegió la población de estas ballenas de una mayor explotación.[83] [84]
Los balleneros del siglo XVIII, se iniciaron con balandras que podían llevar solo uno o dos botes. La cobertura de la flota y tamaño se incrementó con el tiempo, y barcos más grandes entraron al negocio. A finales del siglo XVIII e inicios del XIX los balleneros navegaban por los océanos Pacífico e Índico, el Japón, las costas de Arabia, Australia y Nueva Zelanda.[82] [85] [86] La cacería era peligrosa para la tripulación. Por ejemplo, el 20 de noviembre de 1820, un cachalote que se calculó medía 25,9 metros de longitud atacó al barco ballenero de Nantucket Essex. Solo 8 de los 21 marineros sobrevivieron al ser rescatados por otros barcos.[87]
La cacería declinó desde la década de 1880 hasta 1946, pero se reactivó de nuevo luego de la Segunda Guerra Mundial. Los balleneros modernos eran más eficientes que los que cazaban en botes, empleando barcos a vapor y arpones de explosión. Inicialmente la actividad se centró en las grandes ballenas barbadas misticetos, pero al mermar la población de éstas, la cacería de los cachalotes se incrementó. Los cosméticos, jabones y lubricantes para maquinaria eran los artículos con más compradores. Después que la población disminuyó en forma significativa, la Comisión Ballenera Internacional dio a la especie protección total en 1985. La cacería por por parte del Japón en el norte del océano Pacífico continuó hasta 1988.[84]
Se estima que la población histórica por todo el mundo sumaba 1.100.000 ejemplares antes de la explotación comercial a principios del siglo XVIII.[1] Para 1880 ésta había declinado a un estimado de 29 por ciento de la inicial.[1] Desde ese momento hasta 1946 la población pareció haberse recuperado al disminuir la cacería por parte de los balleneros, pero después de la Segunda Guerra Mundial, disminuyo una vez más, a solo el 33 por ciento de la estimada, antes de la irrupción de la industria ballenera.[1] Se estima que en el siglo XIX fueron capturados entre 184.000 y 236.000 ejemplares,[88] mientras que en la era moderna, al menos 770.000 fueron cazados, la mayoría entre 1946 y 1980. De estos más de 680.000 fueron oficialmente reportados.[89] Revisiones adicionales encontraron que reportes oficiales de la URSS mencionan al menos 89.000 capturas.[90] En otros países como el Japón se encontró faltantes en el registro de capturas.[91]
La población restante de estos animales es lo suficientemente grande para que el estado de conservación de la especie sea considerada como vulnerable, en lugar de en peligro de extinción.[1] Sin embargo, la recuperación de los años de cacería, es un proceso lento, principalmente en el sur del océano Pacífico, donde la cuota de machos en edad reproductiva fue alta.[92]
Estado de conservación actual
El número de cachalotes a través del mundo es desconocido, pero se piensa que es de cientos de miles.[1] El panorama de conservación es más claro que el de muchas otras ballenas. Históricamente Japón ha capturado diez individuos al año y hasta 2006 diez de ellos fueron capturados en Indonesia. Están protegidos prácticamente por todo el mundo y la caza comercial ha cesado.[1] Las criaturas que el cachalote come no son blanco de los pescadores.[1] Sin embargo, los operadores de pesca mediante líneas en el golfo de Alaska se han quejado del robo de pescado de sus líneas de pesca.[52]
Actualmente las principales amenazas que enfrentan estos cetáceos, son los enredones en las líneas de pesca y colisiones con barcos.[11] Otros peligros actuales incluyen la ingestión de desechos marinos, el ruido, y la contaminación química.[93] Para la UICN este cetáceo permanece como «vulnerable».[1] Las lista de los EE. UU. lo ubican como «en peligro de extinción».
Importancia cultural
 Diente de cachalote con un grabado (Scrimshaw).
Diente de cachalote con un grabado (Scrimshaw).Los dientes de cachalote montados en cuerdas son un objeto cultural importante a través del pacífico. En Nueva Zelanda los maorís los conocían como «rei puta» y eran escasos debido a que no eran cazados activamente en la sociedad tradicional maorí.[94] El marfil y huesos eran tomados de ejemplares varados. En Fiyi los dientes eran conocidos como «tabua» y eran tradicionalmente dados como regalo para expiación o muestra de estimación (llamado «sevusevu»), y era importante en negociaciones entre jefes rivales.[95] Friedrich Ratzel en La Historia de la Humanidad reportó en 1896, que en Fiyi, los dientes de cachalote era el artículo más demandado como ornamento o valor. Estos se organizaban a menudo en collares.[96] Hoy la tabua permanece como un elemento importante en la vida de los Fiyianos. Los dientes eran originalmente raros en Fiyi y Tonga, la cual exportaba dientes, pero con la llegada de los europeos, los dientes inundaron el mercado y esta «moneda» colapsó. La sobreoferta permitió a su vez el desarrollo del arte europeo de confección en piezas de ballena (huesos y dientes) llamado en inglés «Scrimshaw».[97]
La ballena de la novela Moby-Dick de Herman Melville es un cachalote.[98] [99] Melville lo asoció con el Leviatán bíblico.[99] [100] La reputación de fiereza perpetuada por Melville, se basó en la habilidad de los cachalote para defenderse con eficacia de la cacería de los primeros balleneros, resultando ocasionalmente en la destrucción de los navíos.
Julio Verne en Veinte mil leguas de viaje submarino, los menciona (quizá incorrectamente) cazando ballenas de su misma especie. En la película de Disney Pinocho de 1940, «Monstro» la ballena es un cachalote mediano muy similar a Moby-Dick excepto que «Monstro» era más negro que blanco.
En 2006 la BBC emitió el programa de televisión titulado Ocean Odyssey (en América Latina Odisea en el Océano), en el cual se recrea la vida de un cachalote con ayuda de imágenes generadas por computador, desde su primera inmersión a los 2 años de edad, hasta su muerte a los 80 años varado en una playa.[101]

Avistamiento de cachalotes
Los cachalotes no son los cetáceos más fáciles de observar, debido al largo tiempo que pasan sumergidos y a su habilidad para atravesar grandes distancias bajo el agua. Sin embargo, debido a su apariencia distintiva y a su gran tamaño se han hecho muy populares. Los observadores a menudo usan hidrófonos para escuchar los chasquidos y así localizarlos antes que estos emerjan. Sitios populares para su observación se encuentran en Kaikoura ubicada en la Isla del Sur de Nueva Zelanda, donde la plataforma continental en tan angosta que las ballenas pueden ser observadas desde la costa, Andenes y Tromsø en la Noruega ártica y en las Islas Azores, donde pueden verse a lo largo del año.[41] [102] Dominica, se cree, es la única isla del Caribe con una población residente de hembras y crías durante todo el año.[103]
Referencias
- ↑ a b c d e f g h i j k Taylor, B.L., Baird, R., Barlow, J., Dawson, S.M., Ford, J., Mead, J.G., Notarbartolo di Sciara, G., Wade, P. y Pitman, R.L. (2008). Physeter macrocephalus. En: UICN 2008. Lista Roja de Especies Amenazadas UICN. Consultado el 7 de octubre de 2008.
- ↑ Haupt, P. (1907). «Jonah's Whale». Proceedings of the American Philosophical Society 46 (185): pp. 151–164. http://www.jstor.org/pss/983449. Consultado el 9 de noviembre de 2008.
- ↑ Corominas, Joan (1987). Breve diccionario etimológico de la lengua castellana. Madrid: Gredos. ISBN 84-249-1 332-9.
- ↑ a b c d e f g h i j k l Shirihai, H. y Jarrett, B. (2006). Whales Dolphins and Other Marine Mammals of the World. Princeton: Princeton Univ. Press. pp. 21–24. ISBN 0-691-12757-3.
- ↑ a b c d e f g h i j k l m n ñ o Whitehead, H. (2002). «Sperm whale Physeter macrocephalus». En Perrin, W., Würsig B. y Thewissen, J.. Encyclopedia of Marine Mammals. Academic Press. pp. 1165–1172. ISBN 0-12-551340-2.
- ↑ «Physeter macrocephalus, Sperm Whale». Consultado el 9 de noviembre de 2009.
- ↑ Shirihai, H. y Jarrett, B. (2006). Whales Dolphins and Other Marine Mammals of the World. Princeton: Princeton Univ. Press. pp. 112–115. ISBN 0-691-12757-3.
- ↑ Maury, M. (1853). Explanations and Sailing Directions to Accompany the Wind and Current Charts. C. Alexander. p. 297. http://books.google.com/books?ct=result&id=DH8TAAAAYAAJ&dq=maury+sperm+whale&ots=GUMZt0D_P_&pg=PA313&lpg=PA313&q=%22sperm+whale%22#PPA297,M1. Consultado el 16 de noviembre de 2008.
- ↑ a b «Sperm Whale». Consultado el 12 de octubre de 2008.
- ↑ Mark Carwardine (1994). On the Trail of the Whale. Chapter 1. Thunder Bay Publishing Co. ISBN 1-899074-00-7.
- ↑ a b c d e Reeves, R., Stewart, B., Clapham, P. & Powell, J. (2003). Guide to Marine Mammals of the World. Nueva York: A.A. Knopf. pp. 240–243. ISBN 0-375-41141-0.
- ↑ «Sperm Whale (Physeter macrocephalus): Species Accounts». Consultado el 12 de octubre de 2008.
- ↑ «Offshore Cetacean Species». CORE. Consultado el 12 de octubre de 2008.
- ↑ «American Cetacean Society Fact Sheet». Consultado el 19 de marzo de 2007.
- ↑ «Sperm Whale Facts». Consultado el 26 de diciembre de 2007.
- ↑ Whitehead, H. (2003). Sperm Whales Social Evolution in the Ocean. Chicago: University of Chicago Press. p. 4. ISBN 0-226-89518-1.
- ↑ «Sperm Whale». Oceana. Consultado el 20 de agosto de 2009.
- ↑ The Southwestern Company: "The Volume Library 1", página 65, 1987, ISBN 0-87197-208-5
- ↑ Kooyman, G. L.& Ponganis, P. J. (octubre de 1998). «The Physiological Basis of Diving to Depth: Birds and Mammals». Annual Review of Physiology 60 (1): pp. 19–32. doi:.
- ↑ Tyack, P., Johnson, M., Aguilar Soto, N., Sturlese, A. & Madsen, P. (18 de octubre de 2006). «Extreme diving of beaked whales». Journal of Experimental Biology 209: pp. 4238–4253. doi:. PMID 17050839. http://jeb.biologists.org/cgi/content/full/209/21/4238.
- ↑ Noren, S. R. & Williams, T. M. (junio de 2000). «Body size and skeletal muscle myoglobin of cetaceans: adaptations for maximizing dive duration». Comparative Biochemistry and Physiology - Part A: Molecular & Integrative Physiology 126 (2): pp. 181–191. doi:.
- ↑ Marshall, C. (2002). «Morphology, Functional; Diving Adaptations of the Cardiovascular System». En Perrin, W., Würsig B. y Thewissen, J.. Encyclopedia of Marine Mammals. Academic Press. p. 770. ISBN 0-12-551340-2.
- ↑ «Aquarium of the Pacific - Sperm Whale». Aquarium of the Pacific. Consultado el 06 de noviembre de 2008.
- ↑ Shwartz, Mark. «Scientists conduct first simultaneous tagging study of deep-diving predator, prey», Stanford Report, 8 de marzo de 2007. Consultado el 6 de noviembre de 2008.
- ↑ a b Clarke, M. (1978). «Structure and Proportions of the Spermaceti Organ in the Sperm Whale». Journal of the Marine Biological Association of the United Kingdom 58: pp. 1–17. http://sabella.mba.ac.uk/2028/01/Structure_and_proportions_of_the_spermaceti_organ_in_the_sperm_whale.pdf. Consultado el 5 de noviembre de 2008.
- ↑ Moore MJ, Early GA (2004). «Cumulative sperm whale bone damage and the bends». Science 306 (5705): pp. 2215. doi:. PMID 15618509.
- ↑ Whitehead, H. (2003). «Foraging». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 156–161. ISBN 0-226-89518-1.
- ↑ «Sperm Whales (Physeter macrocephalus)». U.S. Department of Commerce NOAA Office of Protected Resources. Consultado el 17 de noviembre de 2008.
- ↑ a b Marino, L. (2004). «Cetacean Brain Evolution Multiplication Generates Complexity». International Journal of Comparative Psychology 17: pp. 3–4. http://www.dauphinlibre.be/CetaceanBrainEvolutionIJCP.pdf.
- ↑ Dicke, U., Roth, G.. «Intelligence Evolved», agosto/septiembre de 2008, pp. 71–77.
- ↑ Cranford, T.W. (2000). «In Search of Impulse Sound Sources in Odontocetes». En Au, W.W.L, Popper, A.N. & Fay, R.R.. Hearing by Whales and Dolphins (Springer Handbook of Auditory Research series). Springer-Verlag, Nueva York. ISBN 0387949062.
- ↑ Whitlow, W. (2002). «Echolocation». En Perrin, W., Würsig B. y Thewissen, J.. Encyclopedia of Marine Mammals. Academic Press. pp. 359–367. ISBN 0-12-551340-2.
- ↑ Clarke, M. (1978). «Physical Properties of Spermaceti Oil in the Sperm Whale». Journal of the Marine Biological Association of the United Kingdom 58: pp. 19–26. http://sabella.mba.ac.uk/2029/01/Physical_properties_of_spermaceti_oil_in_the_sperm_whale.pdf. Consultado el 5 de noviembre de 2008.
- ↑ Clarke, M.R. (noviembre de 1970). «Function of the Spermaceti Organ of the Sperm Whale». Nature 228: pp. 873–874. doi:.
- ↑ «Spermaceti as battering ram?» (PDF). Consultado el 19 de marzo de 2007.
- ↑ Carrier, D., Deban, S. & Otterstrom, J. (2002). «The face that sank the Essex: potential function of the spermaceti organ in aggression». The Journal of Experimental Biology 205: pp. 1755–1763. http://autodax.net/Carrieretal2002.pdf.
- ↑ spermwhale.org - Cranford
- ↑ Cranford, T.W. (1999). «The Sperm Whale's Nose: Sexual Selection on a Grand Scale?». Marine Mammal Science 15 (4): pp. 1133–1157. doi:.
- ↑ Whitehead, H. (2003). «Oceanographic Habitat of the Sperm Whale». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. p. 33. ISBN 0-226-89518-1.
- ↑ Murray, J. W., Jannasch, H. W., Honjo, S., Anderson, R. F., Reeburgh, W. S., Top, Z., Friederich, G. E., Codispoti, L. A. & Izdar E. (30 de marzo de 1989). «Unexpected changes in the oxic/anoxic interface in the Black Sea». Nature 338: pp. 411–413. doi:.
- ↑ a b Whitehead, H. (2003). «Sperm Whales and Humans». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 23–24. ISBN 0-226-89518-1.
- ↑ Whitehead, H. & Weilgart, L. (2000). «The Sperm Whale». En Mann, J., Connor, R., Tyack, P. & Whitehead, H.. Cetacean Societies. The University of Chicago Press. p. 169. ISBN 0-226-50341-0.
- ↑ Whitehead, H. (2003). «Mating Systems». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 271–285. ISBN 0-226-89518-1.
- ↑ Pitman, RL; Ballance, LT; Mesnick, SI; Chivers, SJ (2001) "Killer whale predation on sperm whales: Observations and implications" Marine Mammal Science 17(3): 494-507 Abstract
- ↑ Whitehead, H. & Weilgart, L. (2000). «The Sperm Whale». En Mann, J., Connor, R., Tyack, P. & Whitehead, H.. Cetacean Societies. The University of Chicago Press. p. 165. ISBN 0-226-50341-0.
- ↑ Piper, Ross (2007), Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals, Greenwood Press.
- ↑ Estes, J. (2006). Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 179. ISBN 0520248848. http://books.google.com/books?id=daY_utPoJGAC&pg=PA179&lpg=PA179&dq=%22sperm+whale%22+%22killer+whale%22+predator+male&source=web&ots=5F7Z4YEHc_&sig=huE7rZbpMb55EaDcXsUW32Vu0Co&hl=en&sa=X&oi=book_result&resnum=3&ct=result#PPA179,M1. Consultado el 03 de noviembre de 2008.
- ↑ a b Whitehead, H. (2003). «Vertical Movements: The Sperm Whale's Dive». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. p. 79. ISBN 0-226-89518-1.
- ↑ a b Whitehead, H. (2003). «The Diet of a Sperm Whale: The Walnut, the Pea and the Half-Pound Steak». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 43–55. ISBN 0-226-89518-1.
- ↑ Smith S. & Whitehead, H. (2000). «The Diet of Galapagos sperm whales Physeter macrocephalus as indicated by fecal sample analysis». Marine Mammal Science 16 (2): pp. 315–325. doi:.
- ↑ Gaskin D. & Cawthorn M. (1966). «Diet and feeding habits of the sperm whale (Physeter macrocephalus L.) in the Cook Strait region of New Zealand». New Zealand Journal of Marine and Freshwater Research 2: pp. 156–179.
- ↑ a b «Sneaky Cetaceans». Arctic Science Journeys. Consultado el 4 de noviembre de 2008.
- ↑ «Whale Buffet». Consultado el 19 de marzo de 2008.
- ↑ «animal facts: megamouth shark» (en inglés). Consultado el 29 de enero de 2010.
- ↑ «Sperm Whales Spotted Attacking Megamouth Shark» (en inglés). Consultado el 29 de enero de 2010.
- ↑ Compagno, L. J. V. (2001). Sharks of the World Volume 2 Bullhead, mackerel and carpet sharks. FAO Species Catalogue for Fishery Purposes. pp. 74–78. ftp://ftp.fao.org/docrep/fao/009/x9293e/x9293e00.pdf.
- ↑ Dannenfeldt K.H. (1982). «Ambergris: The Search for Its Origin». Isis 73 (3): pp. 382–397. doi:.
- ↑ Ellis, R. (1994). Monsters of the Sea. The Lyons Press. p. 245. ISBN 1592289673.
- ↑ a b c d e Bianucci, G. & Landini, W. (8 de septiembre de 2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society 148 (1): pp. 103–131. doi:.
- ↑ Møhl, B., Wahlberg, M., Madsen, P. T., Heerfordt, A. y Lund, A. (agosto de 2003). «The monopulsed nature of sperm whale clicks». The Journal of the Acoustical Society of America 114 (2): pp. 1143–1153. doi:.
- ↑ Benoit-Bird K. Au W. & Kastelein R. (agosto de 2006). «Testing the odontocete acoustic prey debilitation hypothesis: No stunning results». The Journal of the Acoustical Society of America 120 (2): pp. 1118–1123. doi:.
- ↑ ITIS standar report. «Kogiidae» (en inglés). Consultado el 30 de enero de 2010.
- ↑ a b c d Lambert, O., Bianucci, G. & de Muizon, C. (agosto de 2008). «A new stem-sperm whale (Cetacea, Odontoceti, Physeteroidea) from the Latest Miocene of Peru». Comptes Rendus Palevol 7 (6): pp. 361–369. doi:.
- ↑ Linnaeus, Carolus (1758). Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata.. Holmiae. (Laurentii Salvii).. pp. 824.
- ↑ a b Fordyce, R.E. y Barnes, L.G. (mayo de 1994). «The Evolutionary History of Whales and Dolphins». Annual Review of Earth and Planetary Sciences 22: pp. 419–455. doi:. http://www.saddleback.edu/faculty/thuntley/papers/fordyce_barnes_1994.pdf. Consultado el 4 de octubre de 2008.
- ↑ Stucky, R.E. y McKenna, M.C. (1993). «Mammalia». En Benton, M.J.. The Fossil Record. London.: Chapman & Hall. pp. 739–771.
- ↑ a b c d e f g Mchedlidze. G. A. (2002). «Sperm whales, evolution». En Perrin, W. F., Würsic, B. & Thewissen, J. G. M.. Encyclopedia of Marine Mammals. Academic Press, San Diego. pp. 1172–1174. ISBN 0-12-551340-2.
- ↑ a b Hirota, K. & Barnes, L. G. (5 de abril de 2006). «A new species of Middle Miocene sperm whale of the genus Scaldicetus (Cetacea; Physeteridae) from Shiga-mura, Japan». Island Arc 3 (4): pp. 453–472. doi:.
- ↑ Bianucci, G., Landrini, W. & Varola, W. (septiembre/octubre de 2004). «First discovery of the Miocene northern Atlantic sperm whale Orycterocetus in the Mediterranean». Geobios 37 (5): pp. 569–573. doi:.
- ↑ a b c Nikaido, M., Matsuno, F., Hamilton, H., Brownwell, R., Cao, Y., Ding, W., Zuoyan, Z., Shedlock, A., Fordyce, R. E., Hasegawa, M. & Okada, N. (19 de junio de 2001). «Retroposon analysis of major cetacean lineages: The monophyly of toothed whales and the paraphyly of river dolphins». Procedings of the National Academy of Sciences of the United States of America 98 (13): pp. 7384–7389. doi:. http://www.pnas.org/content/98/13/7384.full. Consultado el 1 de noviembre de 2008.
- ↑ a b Whitehead, H. (2003). «Evolutionary History of the Sperm Whale». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 2–3. ISBN 0-226-89518-1.
- ↑ Heyning, J. (23 de agosto de 2006). «Sperm Whale Phylogeny Revisited: Analysis of the Morphological Evidence». Marine Mammal Science 13 (4): pp. 596–613. doi:.
- ↑ Wilson, D. (1999). The Smithsonian Book of North American Mammals. Vancouver: UBC Press. p. 300. ISBN 0774807628.
- ↑ The Southampton Oceanography Centre & A deFontaubert. «The status of natural resources on the high seas» pág. 63. IUCN. Consultado el 11 de octubre de 2008.
- ↑ Jamieson, A. (1829). A Dictionary of Mechanical Science, Arts, Manufactures, and Miscellaneous Knowledge. H. Fisher, Son & Co.. p. 566.
- ↑ «Aquarium of the Pacific - Sperm Whale». Consultado el 11 de octubre de 2008.
- ↑ Whitehead, H. (2003). «Sperm Whales and Humans». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. p. 14. ISBN 0-226-89518-1.
- ↑ Simons, B.. «Christopher Hussey Blown Out (Up) to Sea». Nantucket Historical Association.
- ↑ Dudley, P. (1725). «An Essay upon the Natural History of Whales, with a Particular Account of the Ambergris Found in the Sperma Ceti Whale». Philosophical Transactions (1683-1775), Vol. 33. The Royal Society. p. 267. http://www.capecodhistory.us/18th/Dudley-whales-1724.htm.
- ↑ a b Dolin, E. (2007). Leviathan: The History of Whaling in America. W. W. Norton. pp. 98–100. ISBN 0393060578.
- ↑ Starbuck, A. (1878). History of the American Whale Fishery from its Earliest Inception to the Year 1876. http://mysite.du.edu/~ttyler/ploughboy/starbuck.htm#sectiond.
- ↑ a b c Bockstoce, J. (diciembre de 1984). «From Davis Strait to Bering Strait: The Arrival of the Commercial Whaling Fleet in North America's West Arctic». Arctic 37 (4): pp. 528–532. http://pubs.aina.ucalgary.ca/arctic/Arctic37-4-528.pdf.
- ↑ Estes, J. (2006). Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 329. ISBN 0520248848.
- ↑ a b Whitehead, H. (2003). «Sperm whales and humans». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 13–21. ISBN 0-226-89518-1.
- ↑ Stackpole, E. A. (1972). Whales & Destiny: The Rivalry between America, France, and Britain for Control of the Southern Whale Fishery, 1785-1825. The University of Massachusetts Press. ISBN 0870231049.
- ↑ Baldwin, R., Gallagher, M. y van Waerebeek, K.. «A Review of Cetaceans from Waters off the Arabian Peninsula» pág. 6. Consultado el 15 de octubre de 2008.
- ↑ «The Wreck of the Whaleship Essex». BBC. Consultado el 11 de octubre de 2008.
- ↑ Davis, L, Gallman, R. & Gleiter, K. (1997). In Pursuit of Leviathan: Technology, Institutions, Productivity, and Profits in American Whaling, 1816-1906 (National Bureau of Economic Research Series on Long-Term Factors in Economic Dev). University of Chicago Press. p. 135. ISBN 0226137899.
- ↑ «Whaling Statistics». Consultado el 15 de octubre de 2008.
- ↑ «Sperm Whale (Physeter macrocephalus) California/Oregon/Washington Stock». Consultado el 16 de octubre de 2008.
- ↑ «The RMS - A Question of Confidence: Manipulations and Falsifications in Whaling». Consultado el 16 de octubre de 2008.
- ↑ Whitehead, H. (2003). «Ghosts of Whaling Past». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 360–362. ISBN 0-226-89518-1.
- ↑ Whitehead, H. (2003). «New Threats to Sperm Whales». Sperm Whales Social Evolution in the Ocean. University of Chicago Press. pp. 362–368. ISBN 0-226-89518-1.
- ↑ «Museum of New Zealand Te Papa Tongarewa Collections Online Search - Rei puta». Consultado el 15 de marzo de 2009.
- ↑ Arno, A. (2005). «Cobo and tabua in Fiji: Two forms of cultural currency in an economy of sentiment». American Ethnologist 32 (1): pp. 46–62. doi:. http://cat.inist.fr/?aModele=afficheN&cpsidt=16581746. Consultado el 6 de octubre de 2008.
- ↑ Ratzel, Friedrich. The History of Mankind. (Londres: MacMillan, 1896). Consultado el 21 de octubre de 2008.
- ↑ Constantine, R. (2002). «Folklore and Legends». En Perrin, W., Würsig, B. & Thewissen, J.. Encyclopedia of Marine Mammals. Academic Press. p. 449. ISBN 0-12-551340-2.
- ↑ «Chapter 3. Romances of Adventure. Section 2. Herman Melville. Van Doren, Carl. 1921. The American Novel». Bartleby.com. Consultado el 19 de octubre de 2009.
- ↑ a b Zwart, H. (2000). What is a Whale? Moby Dick, marine science and the sublime. Tubingen Attempo. 185–214. http://www.filosofie.science.ru.nl/research/hra/whale.pdf.
- ↑ Edwards, B.. «The Playful Learnings». Australasian Journal of American Studies: p. 9. http://www.anzasa.arts.usyd.edu.au/a.j.a.s/Articles/1_06/EdwardsArticle.pdf.
- ↑ Mundo plus TV. «"Odisea en el Océano" se adentra en las maravillas». Consultado el 14 de julio de 2010.
- ↑ «Whale and dolphin watching in the Azores». Wildlife Extra. Consultado el 26 de agosto de 2009.
- ↑ «Whale Watching Dominica». Consultado el 26 de septiembre de 2009.
Enlaces externos
- Physeter en Wikispecies.
- Physeter en "Fauna ibérica; mamíferos". Ángel Cabrera Latorre. 1914. Facsímil electrónico.
- Physeter macrocephalus (como Ph. catodon) en el mismo sitio.
 Wikimedia Commons alberga contenido multimedia sobre Physeter macrocephalusCommons.
Wikimedia Commons alberga contenido multimedia sobre Physeter macrocephalusCommons. Wikiespecies tiene un artículo sobre Physeter macrocephalus. Wikispecies
Wikiespecies tiene un artículo sobre Physeter macrocephalus. Wikispecies- Whale science
- Vidéos de cachalots En francés
- US National Marine Fisheries Service Sperm Whale web page
- 70South - information on the sperm whale
- MarineBio: Sperm Whale, Physeter catodon
- "Physty"-stranded sperm whale nursed back to health and released in 1981
- The physiology of the deep diving adaptations of whales
- ARKive Fotografías, video.
- Whale Trackers - An online documentary film buscando los cachalotes en el mar Mediterráneo.
- El profesor Malcolm Clarke habla sobre la anatomía de los cachalotes Clarke ha pasado gran parte de su vida estudiando los cachalotes y calamares gigantes.
Categorías:- Cetáceos en estado vulnerable
- Physeteridae
- Cetáceos del Atlántico
- Cetáceos del Pacífico
- Cetáceos del Índico











Wikimedia foundation. 2010.