- Teoría cable
-
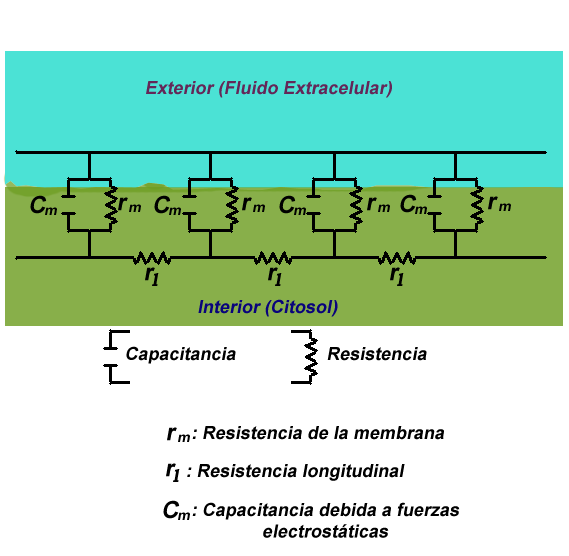
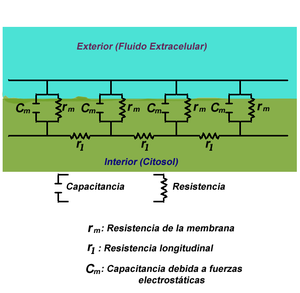 Figura.1: Visión simplificada de la teoría del cable en una fibra neuronal.
Figura.1: Visión simplificada de la teoría del cable en una fibra neuronal.
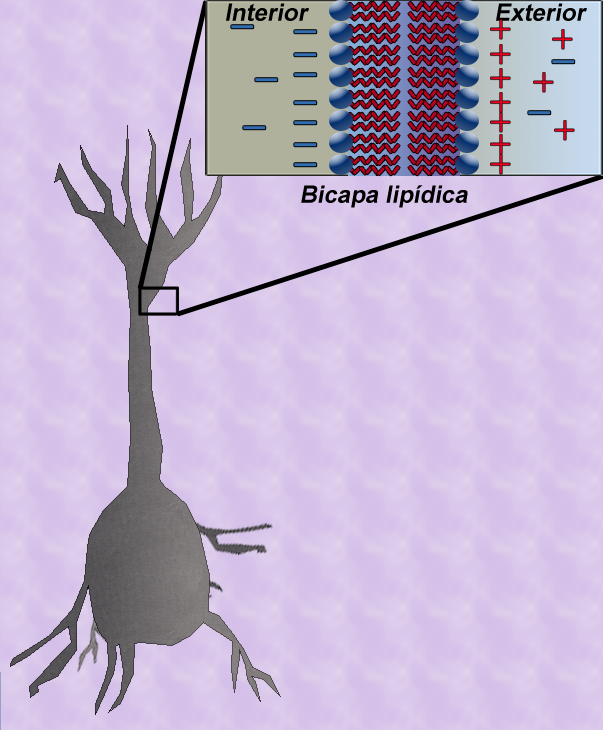
La teoría del cable clásica utiliza modelos matemáticos para calcular el flujo de una corriente eléctrica (y de la tensión asociada) a lo largo de fibras neuronales (axones) pasivas [1] La plantilla {{ref}} está obsoleta, véase el nuevo sistema de referencias., en particular las dendritas que reciben la entrada de las sinapsis en diferentes lugares y tiempos. Las estimaciones se realizadan por el modelado dendritas y axones como cilindros compuestos de segmentos con capacitancia cm y resistencia rm combinadas en paralelo (Ver Figura 1). La capacitancia de una fibra neuronal se produce porque fuerzas electrostáticas están actuando a través de una bicapa lipídica muy delgada (Ver Figura 2). Las resistencias en serie a lo largo de la fibra rl son debidas a la fuerte resistencia que el citosol impone al movimiento de cargas eléctricas.
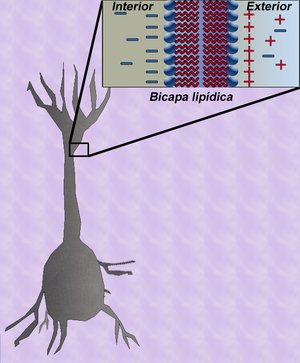 Figure.2: Capacitancia en una fibra.
Figure.2: Capacitancia en una fibra.Contenido
Historia
El origen de la teoría del cable en neurociencia computacional se remonta a la década de 1850, cuando el profesor William Thomson (más tarde conocido como Lord Kelvin) comenzó a desarrollar modelos matemáticos sobre la corrupción de la señal en los cables de telégrafo sumergidos bajo el agua. Los modelos se parecían a las ecuaciones diferenciales parciales utilizadas por Fourier para describir la conducción de calor en un cable.
La década de 1870 vio las primeras aportaciones de Hermann von Helmholtz al modelo electrotónico axónal también enfocado en analogías con la conducción de calor. Sin embargo fue Hoorweg quien descubrió las analogías con los cables submarinos de Kelvin en 1898 y luego Hermann y Cremer de manera independiente, desarrollaron la teoría del cable de fibras neuronales a principios del siglo XX. Otras teorías matemáticas de la conducción de la fibra nerviosa sobre la base de la teoría del cable fueron desarrollados por Cole y Hodgkin (1920 a 1930), Offner et al. (1940), y Rushton (1951).
La evidencia experimental de la importancia de la teoría del cable en el modelado de los axones nerviosos reales comenzó a aparecer en la década de 1930 a partir de los trabajos de Cole, Curtis, Hodgkin, Katz, Rushton, Tasaki y otros. En esta época fueron muy importantes los papeles de Davis y Lorente de No (1947) y los de Hodgkin y Rushton (1946).
En la década de 1950 mejoraron en las técnicas para medir la actividad eléctrica de neuronas individuales. Así, la teoría del cable llegó a ser importante para el análisis de los datos recogidos a partir de grabaciones de microelectrodos intracelulares y para el análisis de las propiedades eléctricas de las dendritas neuronales. Científicos como Coombs, Eccles, Fatt, Frank, Fuortes y otros se basaron en gran medida en la teoría de cables para obtener mayor conocimiento sobre el funcionamiento de las neuronas y para orientarse en el diseño de nuevos experimentos.
Más tarde, la teoría del cable con sus derivados matemáticos permitió modelos neuronales cada vez más sofisticados para ser explorados por investigadores como Jack, Christof Koch, Noble, Poggio, Rall, Redman, Rinzel, Idan Segev, Shepherd, Torre y Tsien. Una importante vertiente de investigación se centró en analizar los efectos de las diferentes distribuciones de entrada sináptica en la superficie dendrítica de una neurona.
Deduciendo la ecuación del cable
Los valores rm y cm introducidos anteriormente se miden en unidades de longitud de fibra (por lo general centímetros (cm)). Así rm se mide en ohmios-centímetros (Ω·cm) y cm en microfaradios por centímetro (µF/cm). Esto contrasta con Rm[O·cm²] y Cm[µF/cm²], que representan la resistencia específica y capacitancia de la membrana medida dentro de una unidad de superficie de membrana (cm2). Así, si el radio a del cable es conocido[2] La plantilla {{ref}} está obsoleta, véase el nuevo sistema de referencias. y por lo tanto, su circunferencia 2πa, rm y cm se pueden calcular como sigue:
 (1)
(1) (2)
(2)
Esto toma sentido, porque cuanto mayor sea la circunferencia del área más grande para la carga que escapa a través de la membrana, menor será la resistencia (se divide Rm por 2pa) y la membrana almacenará más carga (se multiplica Cm por 2pa). De forma similar, La resistencia específica Rl del citoplasma permite la resistencia longitudinal intracelular por unidad de longitud de rl[O·cm-1], se calcula como: (3)
(3)
De nuevo esto es razonable, porque cuanto mayor es el área de la sección transversal (pa²), mayor es el número de rutas para que la corriente fluya a través del citoplasma y la menor resistencia.Para entender mejor cómo la ecuación cable se deduce simplifiquemos la fibra y supongamos que tiene una membrana completamente cerrada (rm es infinito) sin pérdida de corriente con el exterior, y no hay capacitancia (cm = 0). Una corriente inyectada en la fibra [3] La plantilla {{ref}} está obsoleta, véase el nuevo sistema de referencias. en la posición x = 0 se movería a lo largo del interior de la fibra sin cambios. Alejándose desde el punto de inyección y mediante la ley de Ohm (V = IR) se puede calcular la variación de la tensión como:
 (4)
(4)
Si Δx tiende a cero y tiene incrementos infinitesimales de x, lo podemos escribir (4) como:
 (5)
(5)o
 (6)
(6)
rm tiene el mismo efecto que los agujeros en una manguera de jardín. Cuantos más agujeros más agua se escapará hacia el exterior y menos agua llegará a un punto determinado de la manguera. Del mismo modo en las fibras neuronales algunos flujos que viajan longitudinalmente a lo largo del interior de la fibra se escapan a través de la membrana.Si im es el flujo que se escapa a través de la membrana por unidad de longitud (cm), entonces el flujo total que se escapa a lo largo de y unidades debe ser yim. Así, el incremento de flujo en el citoplasma ?il a una distancia ?x desde la posición x=0 puede escribirse como:
 (7)
(7)o usando incrementos continuos infinitesimales:
 (8)
(8)
im puede expresarse con otra fórmula, mediante la inclusión de la capacitancia, esta provocará un flujo de cargas (corriente) hacia el lado del citoplasma de la membrana. Esta corriente es normalmente conocida como corriente de desplazamiento (en este caso se denota ic.) El flujo sólo tendrá lugar siempre y cuando la capacidad de almacenamiento de la membrana no se alcance. ic puede expresarse como:
 (9)
(9)
donde cm es la capacitancia de la membrana y es el cambio de la tensión en el tiempo. La corriente que pasa por la membrana (ir) puede ser expresada como:
es el cambio de la tensión en el tiempo. La corriente que pasa por la membrana (ir) puede ser expresada como: (10)
(10)
y a causa de que im = ir + ic la siguiente ecuación para im puede obtenerse si no se añade corriente adicional mediante un electrodo: (11)
(11)donde
 representa la carga por unidad de longitud de la corriente longitudinal.
representa la carga por unidad de longitud de la corriente longitudinal.Combinando las ecuaciones (6) y (11) obtendremos una primera versión de la ecuación cable:
 (12)
(12)la cual es una ecuación en derivadas parciales (EDP) de segundo orden.
Con una sencilla reorganización de la ecuación (12) (véase más adelante) es posible hacer aparecer dos términos importantes, es decir, la constante de longitud (a veces denominado el espacio constante) denominada λ y la constante de tiempo denominada τ. Las siguientes secciones van enfocadas hacia estos términos.
La constante de longitud
La constante de longitud denotada con el símbolo λ (lambda) es un parámetro que indica hasta qué punto una corriente se extenderá a lo largo por el interior de una neurita y por lo tanto influir en la tensión a lo largo de esa distancia. Cuanto mayor sea λ más lejos llegará el flujo de corriente. La constante de longitud puede ser expresada como:
 (13)
(13)Esta fórmula tiene sentido, porque cuanto mayor sea la resistencia de la membrana (rm) (dando como resultado mayor valor de λ), más corriente quedará dentro del citosol para viajar longitudinalmente a lo largo de la neurita. Cuanto mayor sea la resistencia citosol (rl) (dando como resultado menor valor de λ), más difícil será para la corriente viajar a través del citosol y menos distancia podrá atravesar. Es posible resolver la ecuación (12) llegando a esta otra:
 (14)
(14)Donde V0 es la despolarización en x = 0 (punto de inyección de la corriente), e es la constante exponencial (valor aproximado 2,71828) y Vx es la tensión a una determinada distancia x de x=0. Cuando x = λ entonces
 (15)
(15)y
Vx = V0e − 1 (16)
lo que significa que cuando se mide V a una distancia λ desde el punto en que x = 0 obtendremos
 (17)
(17)Así Vλ es siempre el 36,8 por ciento de V0.
La constante de tiempo
Los neurocientíficos están interesados en saber lo rápido que cambia el potencial de membrana Vm de una neurita en respuesta a las variaciones de corriente que se inyecta en el citosol. La constante de tiempo τ es un índice que ofrece información sobre esto. τ puede calcularse como:
 (18)
(18)esto parece razonable, porque cuanto mayor sea la capacitancia de la membrana (cm), más corriente se necesitará para cargar y descargar una porción de membrana y más durará este proceso. Así se retardará el potencial de membrana (voltaje a través de la membrana) tras la entrada de corriente. Los tiempos de respuesta varían de 1 a 2 milésimas de segundo en las neuronas que esten procesando información y que necesitan una alta precisión temporal de 100 milisegundos o más. Un tiempo de respuesta típico es de unos 20 milisegundos.
La ecuación cable con las constantes de longitud y tiempo
Si multiplicamos la ecuación (12) por rm en ambos lados de la igualdad se obtiene:
 (19)
(19)y sustituyendo λ2 = rm / rl en el lado izquierdo y τ = cmrm en el lado derecho. La ecuación de cable ahora se puede escribir en su forma más conocida:
 (20)
(20)Referencias
- Nelson, Philip Charles (2005). «Capítulo 12: Impulsos nerviosos». En Reverte. Fisica biologica/ Biological Physics: Sobre la nueva tradicion del siglo XX/ About The New Tradiction For The XX Century. (url con vista previa restringida). pp. 545-590. ISBN 84-291-1837-3. http://books.google.es/books?id=aWu02TNspHMC&lpg=PP1&pg=PA545#v=onepage&q=&f=false.
- Methods in Neuronal Modeling, From synapses to networks, edited by Christof Koch and Idan Segev. ISBN 0-262-61071-X
- Biophysics of Computation, Information Processing in Single Neurons, by Christof Koch. ISBN 0-19-518199-9
- Studies from the Rockefeller Institute for Medical Research, by Davis,L.,Jr. and Lorente de No. R (1947). 131: 442-496.
- The electrical constants of a crustacean nerve fibre, by Hodgkin, A.L. and Rushton, W.A.H. (1946). Proc. Roy. Soc. London. B 133: 444-479.
Notas
- Notas:
- 1 ↑ Esta plantilla está obsoleta, véase el nuevo sistema de referencias. Pasivo aquí se refiere a la resistencia de la membrana independiente de la tensión. Sin embargo, experimentos recientes (Stuart y Sakmann 1994) con membranas dendríticas muestran que muchos de estos están equipados con canales iónicos con los que la resistencia de la membrana es dependiente de la tensión. En consecuencia ha habido una necesidad de actualizar la teoría clásica de cable para acomodar el hecho de que la mayoría de las membranas dendríticas no son pasivas.
- 2 ↑ Esta plantilla está obsoleta, véase el nuevo sistema de referencias. La teoría del cable clásica asume que la fibra tiene un radio constante a lo largo de toda su longitud.
- 3 ↑ Esta plantilla está obsoleta, véase el nuevo sistema de referencias. La teoría del cable clásica asume que las entradas (por lo general inyecciones con un microdispositivo) son las corrientes que se pueden resumir como lineales. Esta linealidad no se cumple para los cambios de conductancia de la membrana sináptica.
Categorías:- Neuropsicología
- Neurociencia computacional
- Sistemas Conexionistas
Wikimedia foundation. 2010.