- Callithrix pygmaea
-
 Tití pigmeo
Tití pigmeo
Estado de conservación 
Preocupación menor (UICN 3.1)Clasificación científica Reino: Animalia Filo: Chordata Clase: Mammalia Orden: Primates Suborden: Haplorrhini Infraorden: Simiiformes Parvorden: Platyrrhini Familia: Callitrichidae Subfamilia: Callitrichinae Género: Callithrix Subgénero: Cebuella Especie: C. pygmaea Nombre binomial Callithrix pygmaea
(Spix 1823) Elliot 1913Distribución 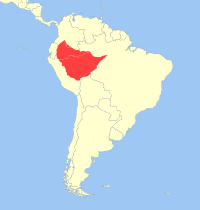
Sinonimia Cebuella pygmaea (Lönnberg, 1940)
Iacchus pygmaeus (Spix, 1823)
Hapale pygmaeus (Schinz, 1825)
Jacchus pygmaeus (Wagner, 1833)
Hapale pygmaea (Wagner, 1848)
Callithrix pygmaeus (Zukowsky, 1937)
Callithrix (Cebuella) pygmaea (Groves, 2001)El tití pigmeo, enano, chichico, mono de bolsillo o de piel roja (Callithrix pygmaea) es una especie de primate platirrino de la subfamilia Callitrichinae. El género Callithrix que son titíes de mandíbula inferior puntiaguda de incisivos tan largos como los colmillos.Es tan pequeño que en la mano de un hombre puede caber una hembra y sus diminutas crías.
Contenido
Taxonomía y filogenía
Cebuella (Gray, 1866) fue propuesto inicialmente como subgénero de Callithrix debido a que están cercanamente relacionados, sin embargo, cuatro años después se consideró como un género distinto conformando entonces un grupo parafilético si el tití pigmeo se seguía considerando (junto con otras especies) dentro de Cebuella, entonces por análisis morfológicos y genéticos se clasificó finalmente bajo el género Callithrix debido a su estrecha relación con el tití amazónico. Es genéticamente cercano a C. argentata difiriendo de C. jacchus por dos características cromosomales.[1]
Existen dentro de Callithrix pygameae dos subespecies basándose en observaciones de campo en el interfluvio de los ríos Purús y Maderia, y especímenes en cautiverio. Se reconoció Callithrix pygmaea niveiventris de pelaje blanquecino en el pecho, abdomen y cara interna de las extremidades ubicado en la orilla sur del Río Solimões; y Callithrix pygmaea pygmaeax donde la superficie ventral es ocre. Sin embargo según otros autores como Hershkovitz[2] exponen argumentos que invalidan el estatus subespecífico dentro de la especie debido a que el color de las extremidades es variable individual y localmente.
Anatomía y fisiología
 Esqueleto de Callithrix pygmaea.
Esqueleto de Callithrix pygmaea.
C. pygmaea es la especie de primate más pequeña del Nuevo Mundo[3] [4] midiendo en etapa adulta 14 a 18 centímetros con una cola no prensil rodeada parcial o completamente de anillos oscuros sobre un fondo claro aún más larga llegando a 15 o 20 centímetros de longitud (casi del 60% del largo total. Los machos más grandes y levemente menos pesados que las hembras (120 y 140 gramos respectivamente),[5] no existe dimorfismo sexual. El pene del adulto no es pigmentado, es liso y cuenta con un báculo de 2,7 mm de longitud y 0,7 de ancho. El escroto es globoso con testículos no pigmentados. Los labios menores femeninos son glandulares, peludos y a veces pigmentados, el clítoris se oculta entre el tejido labial. Las zonas perineal y anal puede ser pigmentada.[2]
Su pelaje, fino, suave y denso, vira de gris a castaño con algunos tonos amarillos y negros con la parte inferior normalmente naranja variando de rojo a blanco. El pelo largo forma una melena que rodea la cara y la nuca casi en su totalidad ocultando las orejas. Tienen las más especializadas y numerosas vibrisas sensoriales de los primates incluyendo conjuntos cubital, carpiano, antebranquial, anconeal, mistacial, mental, interramal, gular y pectoral.[2]
Sus patas traseras son generalmente más largas que las delanteras para su desplazamiento sobre los troncos de árboles. Con garras (tégulas) en todos los dígitos a excepción del dedo gordo del pie en el que tienen una uña plana (ungulae). A diferencia de la mayoría de primates este tiene dos molares y no tres; para una fórmula dental de incisivos 2/2, caninos 1/1, premolares 3/3 y molares 2/2 para un total de 32 dientes.[2] [6] [7] [8] Su cráneo es convexo hacia la parte frontal y el perfil nasal ligeramente cóncavo, las crestas temporales débilmente definidos y con el borde inferior de la mandíbula en forma de V. El volumen de la caja craneal de 19 individuos fue de 34,8 cm3.El sistema esquelético posterior tiene 7 cervicales, 13 torácicas, 6–7 lumbares, 3 S, 28–29 Ca para un total de 57–59 vertebras.[2]
Dieta
Son omnívoros, alimentándose desde savia de específicas especies de plantas en los bosques de galerías a las orillas de ríos y lagunas[3] [9] a frutos e insectos emitiendo sonidos cada vez que una fuente de alimento ha sido encontrada. Realizan agujeros característicos en los árboles absorbiendo la savia, goma y látex y regresando por varios días a nutrirse del mismo orificio. Los agujeros son de 10 a 20 mm de ancho y de 4 a 18 de profundidad siendo hechos primero hacia la base del árbol.[10]
Cuentan además con un ciego en su sistema digestivo que permite aumentar el tiempo de exposición de los fluidos vegetales a los ácidos gástricos. Los caninos inferiores son similares en largo a los incisivos inferiores y los caninos superiores son conspicuos similares a los incisivos adyacentes,[2] condición descrita como procumbente[11] o semiprocumbente[12] posiblemente como una adaptación a la alimentación de exudados.[13] Los incisivos inferiores de C. pygmaea son estrechos y alargados de forma que los cinco dientes en la parte delantera y el centro de la mandíbula inferior son todos la misma longitud permitiendo roer los árboles y estimular el flujo de savia.
Se alimentan de especies vegetales tales como Quararibea rhombifolia (Bombacaceae), Ficus (Moraceae), Pourouma, Vochysia lomatophylla (Vochysiaceae), Trichilia (Meliaceae), Cheiloclinium cognatum (Hippocrataceae) y Spondias mombin (Anacardiaceae).[10] [14] Consumen saltamontes (Orthoptera), abejas silvestres (Hymenoptera), polillas (Lepidoptera), mariposas diurnas (Lepidoptera), libélulas (Odonata) y lagartijas (Anolis, Cnemidophorus lemniscatus, Gonatodes albogularis).[15]
Organización social y comportamiento
Viven en grupo pequeños, de 3 a 8 individuos con un promedio de 5, presumiblemente familias lideradas por una única hembra y un único macho maduros sexualmente, de 0 a 3 subadultos, 0 a 2 juveniles y de 0 a 2 infantes,[16] abarcando el dosel de los árboles que marcan y defienden con glándulas odoríferas del pecho y el área suprapúbica pigmentada o no enmarcada por pelaje negro, y con llamados constantes que incluyen sonidos muy agudos especialmente cuando muestran hostilidad. Las especies de árboles de los que se alimentan varían de grupo a grupo independientemente de la disponibilidad y abundancia dentro de cada una de las localidades.[16]
Ante un posible depredador presentan mobbing, en el que todo el grupo se reúne y atacan al intruso hasta que se retira, o se quedan congelados hasta que la amenaza ha pasado.[14]
Los sitios electos para dormir son nudos densos o raramente huecos en árboles. Son normalmente diurnos empezando actividad un poco después de amanecer. Normalmente se empieza el día con la alimentación (que cuenta con dos picos durante el día, de 6 a 9 y otro de 15 a 18 horas[17] ) para dar paso a actividades sociales como el acicalamiento, el aseo y el juego. Son bastantes ágiles en el movimiento entre los árboles pese a su tamaño, especialmente en las mañanas y atardeceres. Se mueven cuadrúpedamente a lo largo de las ramas saltando hasta cinco metros en las más delgadas. Son de carácter dócil y amable, acicalándose y comunicándose mutuamente con expresiones faciales gracias a la percepción de dos manchas ubicadas a los costados de la boca y una línea blanca vertical hacia las crestas nasales[5] que junto al movimiento amplio de la cabeza, la postura y las vocalizaciones confirman sus relaciones sociales. Este movimiento les permite explorar el ambiente en busca de depredadores mientras se aferran verticalmente a los troncos.
Algunos autores afirman la monogamia en estos grupos, sin embargo, lo observado hasta ahora indica que son grupos poliándricos.[8] La mayoría nacen en dos picos cada año, entre noviembre y enero, y entre mayo y junio. La gestación dura entre dos y tres meses (130 a 150 días) tras los cuales paren en promedio dos crías no idénticas transportadas por los miembros del grupo siendo entonces, todos los adultos partícipes de la crianza y cuidado. El pelaje de los recién nacidos es distinto al de los adultos, con un color amarillo limón que se perderá tras el primer mes. La madre los amamanta hasta en promedio, los tres meses de edad, alcanzando la madurez reproductiva en hembras y machos cerca del año o año y medio y viviendo hasta 11 años en estado silvestre.
Ontogenia y reproducción
La reproducción en estado silvestre se ha observado aproximadamente cada 6 meses. La gestación en cautiverio dura entre 19 y 20 semanas. La edad mínima del primer parto en cautividad es de 32 meses con una frecuencia de casi el 67 al 76% de los nacimientos corresponden a gemelos.[14] [18]
El recién nacido cuenta con pelaje a excepción de su vientre y sus ojos están abiertos al nacer. El desarrollo de los jóvenes se han clasificado en los siguientes grupos de edad:[14] infante 1 (menor de 1 mes) de pequeño tamaño con dientes de leche y llevado por adultos o juveniles; infantil 2 (de 2 a 5 meses) pequeños con locomoción independiente, comienzan a nacer los dientes permanentes; menores (de 6 a 12 meses), de menor tamaño que un adulto y con genitales pequeños, el círculo negro anogenital comienza a aparecer, para finales del noveno mes todos los dientes permanentes han hecho erupción; subadultos (de 12 a 16 meses), siguen siendo ligeramente más pequeños que un adulto con una pigmentación escrotal incipiente y caninos pequeños; a partir de los 16 meses madura sexualmente y los caninos alcanzan la longitud normal.
La erupción dental y la secuencia de desarrollo es similar a otros Callithricidos (mandíbula 5 m1-i1-m2-i2-p3-p2-p1-c1; maxilar 5 M1-I1-M2-(P3–P2)-(P1–I2)-C1), a excepción del hecho que el segundo incisivo entra en erupción después de que el segundo molar.[19] [19] [20] Este retraso en la erupción y el desarrollo de los incisivos es único entre los primates existentes y puede ser correlacionado con la morfología de la corona de los caniniformes. .[21]
Características genotípicas
Callithrix pygmaea tienen 44 cromosomas diploides. El cromosoma Y es acrocéntrico mientras que el X es metacéntrico.[22] [23] El resto de cromosomas (32), los autosomas, son categorizados en 4 grupos basados en el radio y tamaño de los brazos. Se observa que la mayoría son submetacéntricos y 10 son acrocéntricos. La diferencia en el número de cromosomas con su ancestro puede ser causa de la separación ocurrida antes de la diferenciación de los Callithricidae.[24]
A través de la fusión de dos pares de cromosomas acrocéntricos pequeños de Callithrix jacchus, el cariotipo de C. pygmaea pudo haber surgido.[25] Con Callithrix argenata comparte el número de cromosomas (2n=44), mientras que Callithrix jacchus'' tiene el mismo número diploide (2n=46) como Saguinus y Leontopithecus.[26]
La transferencia de células germinales entre los mellizos es posible gracias a la interconexión de sus placentas vía vasos sanguíneos.[27] Por la ocurrencia de dichos partos múltiples en esta especie se observan quimeras XX/XY por anastomosis placentaria con importantes efectos sobre la fertilidad.[28]
Distribución: Poblaciones silvestres
Hábitat y ecología
C. pygmaea es especialista en cuanto al hábitat demostrando preferencia por la vegetación secundaria cercana a los cursos de agua. Habita las tierras bajas tropicales de los bosques de la Amazonía occidental. Su distribución y densidad se correlacionan directamente con la presencia y abundancia de especies de árboles y animales ya mencionados que hacen parte de su dieta.
Habitan selvas lluviosas de poca precipitación o bosques maduros compuestos por cañas, hierbas altas, enredaderas, arbustos y árboles jóvenes hasta los 30 o 40 m en los bordes de ríos en llanuras aluviales inundables periódicamente y por más de tres meses, utilizan los estratos verticales desde el suelo hasta los 20 m. Los datos sobre la precipitación y la temperatura se han establecido sobre un sitio de estudio en el Perú en la cuenca del Río Maniti donde la temporada de lluvias dura de octubre a mayo y la estación seca se extiende de junio a septiembre. La temperatura se mantiene contante a 27 °C y con un 80% de humedad.[29] [30] Los ríos comienzan a aumentar en el comienzo de la temporada seca de tal manera que entre febrero y junio la llanura se inunda.
Los rangos de hogar suelen incluir casi 20 a 60 m tierra adentro y varían en tamaño con una media de media hectárea. Dentro de ella, el tamaño poblacional se ve influido por la densidad de depredadores y competidores, especialmente las hormigas arbóreas que invaden los agujeros hechos por este tití para la alimentación con exudados. Además pueden competir con Saguinus fuscicollis, Saguinus mystax, Callicebus moloch, Saimiri sciureus y Aotus.[14] Los depredadores de C. pygmaea incluyen el capuchino (Cebus apella), el ocelote (Leopardus pardalis), el tigrillo (Leopardus tigrinus y Leopardus wiedii), el puma (Herpailurus yaguarondi), algunas aves de presa (que despiertan llamados de atención cuando vuelan sobre el grupo) y grandes serpientes como la víbora (Bothrops atrox).
Adicionalmente se encuentran vinculados a estos animales algunos parásitos dentro de los que se encuentran nemátodos como Strongyloides stercoralis, Pithecostrongylus alatus, Filaroides cebuellae, Enterobius vermicularis, Primatubulura jacchis y Protospirula sp.; y acantocéfalos como Prosthenorchis elegans y Prosthenorchis spirula.[31]
Distribución actual
Este tití es natural de América del Sur y se distribuyen en la cuenca occidental del Amazonas, al sur del Río Caquetá (Japurá) de Colombia, al oeste en la Amazonía de Ecuador, al sur a través de las tierras bajas del oriente del Perú a la orilla derecha del Río Ucayali, al norte de Bolivia, con una extensión sobre el norte del Río Purús y el Ríoi Solimões de Brasil.[32] [14] [33] [2]
Se reconocen dos subespecies separadas por barreras geográficas como los grandes ríos. Callithrix pygmaea pygmaea es encontrado en el Amazonas, Brasil (al norte del Río Solimões), al este de Perú (al norte del Río Marañón), al sur de Colombia, al norte de Bolivia y noreste de Ecuador. Callithrix pygmaea niveiventris se encuentra en el Amazonas, el Brasil, al este de Perú y al sur y norte de Bolivia.
Estado de conservación y amenazas
La conservación de los primates es fundamental para mantener la estructura y dinámica de los ecosistemas tropicales debido a que distintas especies cumplen roles ecológicos diversos e importantes tales como la de polinización, dispersión de semillas y como eslabón importante en la cadena trófica.[3] Dicha conservación se corresponde directamente con las relaciones establecidas entre los individuos y su espacio, y sus características físicas, comportamentales y poblacionales de cada especie,[34] por lo que es de suma importancia el conocimiento de estos aspectos en las diferentes especies.
Considerando su área de distribución y los problemas como la desaparición de bosques por la ganadería y la agricultura junto con el tráfico ilegal como mascotas hacen que las poblaciones silvestres de esta especie se vean seriamente afectadas, disminuyendo rápidamente,[35] futuro no prometedor a menos de que sean iniciados programas de conservación de los individuos en estado silvestre y de eliminación de las vías de tráfico animal en nuestras selvas Amazónicas.
Vulnerabilidad y categoría de amenaza
La preferencia por determinadas sustancias para su alimentación es de gran importancia ecológica en la repartición de recursos interespecíficos debido a que prefieren alimentos que otros primates evitan.[36] Esta especialización en la dieta, junto con otras características de importancia como las ya mencionadas en cuanto al hábitat, hacen que este primate sea particularmente vulnerable a las actividades humanas, estando ausente en ambientes alterados o en baja densidad poblacional y con cambios comportamentales en áreas con presencia humana como lo son las turísticas; sin embargo, debido a su capacidad de adaptación a bosques secundarios, actualmente se clasifica, según la condición jurídica y social, como una especie de preocupación menor (LC) en la lista roja de la UICN de 2007.[27]
Tratados para su conservación
Esta especie está incluida en el Apéndice II de la Convención sobre el Comercio Internacional de Especies Amenazadas (CITES), lo que significa que el comercio internacional de esta especie debe ser monitorizada cuidadosamente.
Para reducir la degradación del hábitat y las perturbaciones humanas, ha habido algunos esfuerzos para crear conciencia en las comunidades locales. Por ejemplo, un programa en Ecuador se inició para educar a los niños en la importancia de la conservación de los primates.[37]
Tráfico ilegal
Por su diminuto tamaño y bajos niveles de agresividad estos titíes son capturados como mascotas. Los métodos varían pero el más popular es inmovilizar al animal al tronco del que se alimenta con productos químicos. Esta captura afecta dramáticamente el tamaño del grupo y el comportamiento pues se disminuyen los tiempos de juego e interacciones sociales, evitan usar los estratos bajos del bosque y tienen efectos negativos sobre la tasa de reproducción llevando a la desaparición de los grupos en un área. El destino de los animales capturados es también negativo pues muchos de ellos mueren durante los primeros dos meses después de su captura.[cita requerida]
Destrucción de su hábitat
Además de su sensibilidad al aumento en el ruido ambiental y el tráfico humano, otro problema que afecta las poblaciones de esta especie es la destrucción de los bosques de galería en donde los grupos migran o desaparecen debido al agotamiento de las fuentes viejas de exudados y a la ausencia de nuevas fuentes. Por ello, es de importancia proteger las especies de plantas proveedoras de exudados para asegurar la supervivencia de estos primates y de muchas otras especies de animales que también usan esta fuente de alimento.
Referencias
- ↑ BARROSO CML, et al. (1997) Update on the phylogenetic systematics of New World monkeys: further DNA evidence for placing the pygmy marmoset (Cebuella) within the genus Callithrix. International Journal of Primatology 18:651–674.
- ↑ a b c d e f g HERSHKOVITZ P (1977) Living New World monkeys (Platyrrhini), with an introduction to primates. University of Chicago Press, Illinois 1:1–1117.
- ↑ a b c DE LA TORRE S (2000) Primates of Amazonian Ecuador. Proyecto PETRAMAZ/SIMBIOE, Quito.
- ↑ TIRIRA D (2004) Nombres de los mamíferos del Ecuador. Ediciones Murciélago Blanco y Museo Ecuatoriano de Ciencias Naturales. Publicación Especial 5. Quito.
- ↑ a b CAWTHON KA (2005) Primate Factsheets: Pygmy marmoset (Callithrix pygmaea) . Primate Info Net, University of Wisconsin, Madison. Consultado el 12 de noviembre de 2010 [Disponible en http://pin.primate.wisc.edu/factsheets/entry/pygmy_marmoset].
- ↑ MACDONALD D (1984) Encyclopedia of Mammals. Facts on File Publications, New York.
- ↑ RATHBUN GB & FONS R (1990) "Grzimek's Encyclopedia of Mammals". Vol. 1. McGraw-Hill Publishing Co., New York.
- ↑ a b NOWAK RM (1991) "Walker's Mammals of the World". Fifth Edition. Johns Hopkins University Press, Baltimore.
- ↑ DE LA TORRE S, YÉPEZ P & SNOWDON C. en prensa. "Conservation status of pygmy marmosets Callithrix (Cebuella) pygmaea in Amazonian Ecuador". En: The Smallest Anthropoids: the marmoset and callimico radiation. R. Tuttle (ed). Springer Verlag.
- ↑ a b RAMIREZ M, FREESE CH & REVILLA J (1977) "Feeding ecology of the pygmy marmoset, Cebuella pygmaea in northeastern Peru". The biology and conservation of the Callitrichidae (D. G. Kleiman, ed.):91-104. Smithsonian Institution Press, Washington, D.C.
- ↑ MOYNIHAN M (1976) "The New World primates". Princeton University Press, New Jersey.
- ↑ KINZEY WG, ROSENBERGER AL & RAMIREZ M (1975) "Vertical clinging and leaping in a Neotropical anthropoid". Nature 255:327–328.
- ↑ FILHO AF & MITTERMEIER RA (1977) "Tree gouging, exudate-eating and the ‘‘short-tusked’’ condition in Callithrix and Cebuella". The biology and conservation of the Callitrichidae (D. Kleiman, ed.):105-115. Smithsonian Institution Press, Washington, D.C.
- ↑ a b c d e f SOINI P (1988) "The pygmy marmoset, genus Cebuella". Ecology and behavior of Neotropical primates (R. A. Mittermeier, A. B. Rylands, A. Coimbra-Filho, and G. A. B. Fonseca, eds.):79-130. World Wildlife Fund, Washington, D.C.
- ↑ SNOWDON CT & POLA YV (1982) "Prey capture by naive pygmy marmosets (Cebuella pygmaea) ". Laboratory Primate Newsletter 21:4–7.
- ↑ a b DE LA TORRE S, YÉPEZ P, SNOWDON C & PAYAGUAJE A (2004) "Ecología, comportamiento y conservación del leoncillo Callithrix (Cebuella) pygmaea en tierras Secoya". Quito.
- ↑ YÉPEZ P, DE LA TORRE S & SNOWDON CT (2005) "Interpopulation differences in exudate feeding of pygmy marmosets in Ecuadorian Amazonia". American Journal of Primatology 66: 145-158.
- ↑ ZIEGLER TE, SNOWDON CT & BRIDSON WE (1990) "Reproductive performance and excretion of urinary estrogens and gonadotropins in the female pygmy marmoset (Cebuella pygmaea)". American Journal of Primatology 22:191–203.
- ↑ a b BYRD KE (1978) Radiographic analysis of dental development and eruption sequences in the pygmy marmoset, Cebuella pygmaea (Spix 1823). American Journal of Physical Anthropology 48:384.
- ↑ SOINI P & DE SOINI M (1990) "Desarollo dentario y la estimación de la edad en Cebuella pygmaea, Saguinus fuscicollis,y Saguinus mystax". Primatología en el Perú: investigaciones primatológicas 1973–1985:254-271. Proyecto Peruano de Primatología, Lima.
- ↑ TOWNSEND WR (2001) "Callithrix pygmaea. American Society of Mammalogists". Mammalian species 665; 1-6.
- ↑ BENIRSCHKE K & BROWNHILL LE (1962) Further observations on marrow chimerism in marmosets. Cytogenetics 1:245–257.
- ↑ EGOZCUE J, CHIARELLI B & SARTI-CHIARELLI M (1968) "The somatic and meiotic chromosomes of Cebuella pygmaea (Spix, 1823) with special reference to the behavior of the sex chromosome during spermatogenesis". Folia Primatalogica 8:50–57.
- ↑ DE BOER LEM (1974) "Cytotaxonomy of the Platyrrhini (Primates)". Genen en Pnaenen 17:1–115.
- ↑ WOHNUS JF & BENIRSCHKE K (1966) "Chromosome analysis of four species of marmosets (Callithrix jacchus, Tamarinus mystax, Tamarinus nigricollis and Cebuella pygmaea)". Cytogenetics 5:94–105.
- ↑ SEUANEZ HN, FOREMAN L & ALVES G (1988) "Comparative chromosome morphology in three callitrichid genera: Cebuella, Callithrix, and Leontopithecus". Journal of Heredity 79:418–424.
- ↑ a b WISLOCKI GB (1939) "Observations on twinning in marmosets". American Journal of Anatomy 64:445–483.
- ↑ BUENO ML (2003) Importancia de la caracterización genética de especies silvestres en Zoológicos, Unidades de Rescate de fauna y Centros de acopio. Lyonia 3(1):45-56.
- ↑ SOINI P (1982) "Ecology and population dynamics of the pygmy marmoset, Cebuella pygmaea". Folia Primatologica 39:1–21.
- ↑ SOINI P (1986). "A synecological study of a primate community in the Pacaya Samiria Reserve, Peru". Primate Conservation. (7): 63-71.
- ↑ Hershkovitz, P (1977) (en Inglés). Living New World monkeys (Platyrrhini): with an introduction to Primates. Living New World monkeys (Platyrrhini): with an introduction to Primates.
- ↑ JANSON CH & EMMONS LH (1990) "Ecological structure of the nonflying mammal community at Cocha Cashu Biological Station, Manu National Park, Peru". Four Neotropical forests (A. H. Gentry, ed.):314-338. Yale University Press, New Haven, Connecticut.
- ↑ FREESE CW, HELTNE PG, CASTRO N & WHITESIDES G (1982) "Patterns and determinants of monkey densities in Peru and Bolivia, with notes on distributions". International Journal of Primatology 3:53–90.
- ↑ BROCKELMAN R & ALÍ R (1987) Methods of surveying and sampling forest primate population. C. W. Marash y R. A. Mittermeier (ed): Primate Conservation in Tropical Rain Forest. New York;23-63
- ↑ DEFLER T (2003) Primates de Colombia. Serie de guías tropicales de campo. Conservación Internacional Colombia. Bogotá D. C.
- ↑ GLASER D & HOBI G (1985) "Taste responses in primates to citric and acetic acid". International Journal of Primatology 6: 395–398.
- ↑ http://www.arkive.org/pygmy-marmoset/cebuella-pygmaea/ Consultado el 12 de noviembre de 2010.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Callithrix pygmaea. Commons
Wikimedia Commons alberga contenido multimedia sobre Callithrix pygmaea. Commons Wikiespecies tiene un artículo sobre Callithrix pygmaea. Wikispecies
Wikiespecies tiene un artículo sobre Callithrix pygmaea. Wikispecies- Video, Pictures and Information on Pygmy Marmosets
- Primate Info Net Callithrix pygmaea Factsheet
- Nacimiento de gemelos albinos tití pigmeo (Ingles) National Geographic. Photo in the News: Albino Pygmy Monkey Twins Born
- Arkive: Images of life on earth
- Callithrix pygmaea (Spix, 1823)
- CITES (June, 2010)
- IUCN Red List (June, 2007) Cebuella pygmaea (Pygmy marmoset)
- Primate Info Net. Library and Information Service. National Primate Research Center, University of Wisconsin – Madison
Categorías:- Primates no amenazados
- Callitrichinae
- Mamíferos del Amazonas




Wikimedia foundation. 2010.