- Virus
-
 Virus
Virus
Virus de la influenzaClasificación científica Grupos En biología, un virus[n. 1] (del latín virus, «toxina» o «veneno») es una entidad infecciosa microscópica que sólo puede multiplicarse dentro de las células de otros organismos. Los virus infectan todos los tipos de organismos, desde animales y plantas, hasta bacterias y arqueas. Los virus son demasiado pequeños para poder ser observados con la ayuda de un microscopio óptico, por lo que se dice que son submicroscópicos. El primer virus conocido, el virus del mosaico del tabaco,[n. 2] fue descubierto por Martinus Beijerinck en 1899,[1] [2] y actualmente se han descrito más de 5.000, si bien algunos autores opinan que podrían existir millones de tipos diferentes.[3] [4] Los virus se hallan en casi todos los ecosistemas de la Tierra y son el tipo de entidad biológica más abundante.[4] [5] El estudio de los virus recibe el nombre de virología,[6] una rama de la microbiología.[7] [8]
A diferencia de los priones y viroides, los virus se componen de dos o tres partes: su material genético, que porta la información hereditaria, que puede ser ADN o de ARN; una cubierta proteica que protege a estos genes —llamada cápside— y en algunos también se puede encontrar una bicapa lipídica que los rodea cuando se encuentran fuera de la célula —denominada envoltura vírica—. Los virus varían en su forma, desde simples helicoides o icosaedros hasta estructuras más complejas. El origen evolutivo de los virus aún es incierto, algunos podrían haber evolucionado a partir de plásmidos (fragmentos de ADN que se mueven entre las células), mientras que otros podrían haberse originado desde bacterias. Además, desde el punto de vista de la evolución de otras especies, los virus son un medio importante de transferencia horizontal de genes, la cual incrementa la diversidad genética.[9]
Los virus se diseminan de muchas maneras diferentes y cada tipo de virus tiene un método distinto de transmisión. Entre estos métodos se encuentran los vectores de transmisión, que son otros organismos que los transmiten entre portadores. Los virus vegetales se propagan frecuentemente por insectos que se alimentan de su savia, como los áfidos, mientras que los virus animales se suelen propagar por medio de insectos hematófagos. Por otro lado, otros virus no precisan de vectores: el virus de la gripe (rinovirus) se propaga por el aire a través de los estornudos y la tos y los norovirus son transmitidos por vía fecal-oral, o a través de las manos, alimentos y agua contaminados. Los rotavirus se extienden a menudo por contacto directo con niños infectados. El VIH es uno de los muchos virus que se transmiten por contacto sexual o por exposición con sangre infectada.[10]
No todos los virus provocan enfermedades, ya que muchos virus se reproducen sin causar ningún daño al organismo infectado. Algunos virus como el VIH pueden producir infecciones permanentes o crónicas cuando el virus continúa replicándose en el cuerpo evadiendo los mecanismos de defensa del huésped.[11] [12] [13] En los animales, sin embargo, es frecuente que las infecciones víricas produzcan una respuesta inmunitaria que confiere una inmunidad permanente a la infección. Los microorganismos como las bacterias también tienen defensas contra las infecciones víricas, conocidas como sistemas de restricción-modificación. Los antibióticos no tienen efecto sobre los virus, pero se han desarrollado medicamentos antivirales para tratar infecciones potencialmente mortales.[14]
Contenido
Etimología
La palabra proviene del latín virus, que hace referencia al veneno o alguna sustancia nociva, y fue usada por primera vez en inglés en 1392.[15] Virulento, del latín virulentus (venenoso), data del 1400.[16] La mención de «agente que causa enfermedades infecciosa» se usó por primera vez en 1728,[15] antes del descubrimiento de los virus por Dmitri Ivanovski en 1892. El adjetivo "viral" data de 1948.[17] El término «virión» también se utiliza para referirse a una única partícula vírica infecciosa. El plural es «virus».
Historia
 Martinus Beijerinck en su laboratorio en 1921.
Martinus Beijerinck en su laboratorio en 1921.
A mediados del siglo XIX, Louis Pasteur propuso la teoría germinal de las enfermedades, en la cual explicaba que todas las enfermedades eran causadas y propagadas por algún «tipo de vida diminuta» que se multiplicaba en el organismo enfermo, pasaba de éste a otro y lo hacía enfermar.[18] Pasteur, sin embargo, se encontraba trabajando con la hidrofobia, y descubrió que aunque la enfermedad fuera contagiosa y ésta se contrayera por el mordisco de un animal rabioso, no se veía el germen por ningún lado. Pasteur concluyó que el germen sí se encontraba ahí, pero era demasiado pequeño como para poder observarse.[19]
En 1884, el microbiólogo francés Charles Chamberland inventó un filtro (conocido actualmente como filtro Chamberland o filtro Chamberland-Pasteur) que tiene poros de tamaño inferior a los de una bacteria. Así pues, podía hacer pasar por el filtro una solución con bacterias y eliminarlas completamente de la misma.[20] El biólogo ruso Dmitri Ivanovski utilizó este filtro para estudiar lo que actualmente se conoce como virus del mosaico del tabaco. Sus experimentos demostraron que los extractos de hojas molidas de plantas infectadas de tabaco seguían siendo infecciosos después de filtrarlos. Ivanovski sugirió que la infección podría ser causada por una toxina producida por las bacterias, pero no continuó apoyando esta idea.[21] En aquella época se pensaba que todos los agentes infecciosos podían ser retenidos por filtros y, además, que podían ser cultivados en un medio con nutrientes —esta opinión formaba parte de la teoría germinal de las enfermedades.[1] En 1899, el microbiólogo neerlandés Martinus Beijerinck repitió los experimentos de Ivanovski y quedó convencido de que se trataba de una nueva forma de agente infeccioso.[22] Observó que el agente sólo se multiplicaba dentro de células vivas en división, pero como sus experimentos no mostraban que estuviera compuesto de partículas, lo llamó contagium vivum fluidum («germen viviente soluble») y reintrodujo el término «virus».[21] Beijerinck mantenía que los virus eran de naturaleza líquida, una teoría más tarde descartada por Wendell Stanley, que demostró que eran particulados.[21] En ese mismo año, en 1899, Friedrich Loeffler y Frosch pasaron el agente de la fiebre aftosa —el aftovirus— por un filtro similar y descartaron la posibilidad de que se tratara de una toxina debido a la baja concentración, y llegaron a la conclusión de que el agente se podía replicar.[21]
A principios del siglo XX, el bacteriólogo inglés Frederick Twort descubrió los virus que infectaban bacterias, que actualmente se denominan bacteriófagos,[23] y el microbiólogo franco Félix de Herelle describió virus que, cuando se los añadía a bacterias cultivadas en agar, producían zonas de bacterias muertas. Diluyó con precisión una suspensión de estos virus y descubrió que las diluciones más altas, en lugar de matar todas las bacterias, formaban zonas discretas de organismos muertos. Contando estas zonas, y multiplicándoseles por el factor de dilución, De Herelle pudo calcular el número de virus en dicha zona.[24]
A finales del siglo XIX, los virus se definían en términos de su infectividad, filtrabilidad, y su necesidad de huéspedes vivientes. Los virus sólo habían sido cultivados en plantas y animales. En 1906, Ross Granville Harrison inventó un método para cultivar tejidos en linfa, y, en 1913, E. Steinhardt y colaboradores utilizaron este método para cultivar virus Vaccinia en fragmentos de córnea de cobaya.[25] En 1928, H. B. Maitland y M. C. Maitland cultivaron el mismo virus en suspensiones de riñones picados de gallina. Su método no fue adoptado ampliamente hasta 1950, cuando se empezó a cultivar poliovirus a gran escala para la producción de vacunas.[26]
Otro avance se produjo en 1931, cuando el patólogo estadounidense Ernest William Goodpasture cultivó el virus de la gripe y otros virus en huevos fertilizados de gallina.[27] En 1949, John Franklin Enders, Thomas Weller y Frederick Robbins cultivaron virus de la polio en células cultivadas de embriones humanos, siendo la primera vez que se cultivó un virus sin utilizar tejidos animales sólidos o huevos. Este trabajo permitió a Jonas Salk crear una vacuna efectiva contra la polio.[28]
Con la invención de la microscopía electrónica en 1931 por parte de los ingenieros alemanes Ernst Ruska y Max Knoll, se obtuvieron las primeras imágenes de virus.[29] [30] En 1935, el bioquímico y virólogo estadounidense Wendell Stanley examinó el virus del mosaico del tabaco y descubrió que estaba compuesto principalmente de proteínas.[31] Poco tiempo después, el virus fue separado en sus partes de proteínas y de ARN.[32] El virus del mosaico del tabaco fue uno de los primeros en ser cristalizados, y por tanto, la primera estructura que pudo ser observada en detalle. Las primeras imágenes por difracción de rayos X del virus cristalizado fueron obtenidas por Bernal y Fankuchen en 1941. Basándose en sus imágenes, Rosalind Franklin descubrió la estructura completa del virus en 1955.[33] El mismo año, Heinz Fraenkel-Conrat y Robley Williams demostraron que el ARN purificado del virus del mosaico del tabaco y sus proteínas de envoltura pueden reproducirse por sí solos, formando virus funcionales, sugiriendo que este mecanismo sencillo era probablemente como se reproducían los virus en las células huéspedes.[34]
La segunda mitad del siglo XX fue la edad dorada del descubrimiento de virus, y la mayoría de las 2.000 especies reconocidas de virus animales, vegetales y bacterianos fueron descubiertas durante estos años.[35] En 1957, se descubrieron el arterivirus equino y la causa de la diarrea vírica bovina (un pestivirus). En 1963, el virus de la hepatitis B fue descubierto por Baruch Blumberg,[36] y en 1965, Howard Temin describió el primer retrovirus. La transcriptasa inversa, enzima clave que utilizan los retrovirus para convertir su ARN en ADN, fue descrita originalmente en 1970, de manera independiente por Howard Temin y David Baltimore.[37] En 1983, el equipo de Luc Montagnier del Instituto Pasteur de Francia aisló por primera vez el retrovirus actualmente llamado VIH.[38]
Origen
Los virus son organismos parásitos que infectan células y producen viriones para difundir sus genes. La mayoría de las proteínas virales no tienen homólogos en las células modernas, en contradicción con la visión tradicional de los virus como los «ladrones de genes celulares». Esto sugiere que los genes virales básicamente tienen su origen durante la replicación de los genomas virales y/o fueron reclutados de linajes celulares ahora extintos. Algunas proteínas virales específicas están presentes en virus que infectan a los miembros de los tres dominios de la vida,[39] lo que sugiere que los virus son en realidad muy antiguos. En particular, los análisis estructurales de proteínas de la cápside han revelado que al menos dos tipos de viriones se originaron de manera independiente antes que LUCA (el último antepasado universal celular). Aunque varias hipótesis han sido recientemente propuestas para explicar el origen de los virus, la aparición de viriones, como un mecanismo específico para la difusión de genes, permanece sin explicación.
Los avances en la caracterización de los virus a nivel molecular, sugieren que los virus coevolucionan con sus organismos huéspedes,[40] debido a que los virus son parásitos intracelulares extremos y, por lo tanto, requieren de la supervivencia del huésped para poder asegurar su propia supervivencia. Es interesante notar que cuando un virus se replica en su huésped natural, tiende a no causar enfermedad en el mismo o causa una enfermedad leve y autolimitada en la mayoría de los casos. Varios de los virus conocidos producen enfermedades severas sólo cuando infectan organismos diferentes a sus huéspedes naturales. Lo anterior sugiere que buena parte de los virus asociados con la producción de enfermedades, son virus que están en proceso de adaptarse a un nuevo tipo de huésped y que una vez lograda dicha adaptación, la estrategia del virus consiste en perpetuarse y propagarse sin afectar al organismo huésped.
Se pueden encontrar virus dondequiera que haya organismos vivos, y probablemente existen desde la aparición de las primeras células. Su origen es incierto, puesto que no fosilizan, de manera que sólo se puede especular a partir de diferentes técnicas y ensayos de biología molecular.[41] [42] Estas técnicas dependen de la disponibilidad de ADN o ARN vírico antiguo, pero desgraciadamente la mayoría de virus que han sido preservados y almacenados en laboratorios tienen menos de 90 años.[43] [44] Hay tres teorías principales sobre el origen de los virus:[45] [46]
- Teoría de la regresión: es posible que los virus fueran pequeñas células que parasitaban células más grandes. A lo largo del tiempo, los genes que no necesitaban por su parasitismo desaparecieron. Las bacterias Rickettsia y Chlamydia son células vivientes que, como los virus, sólo pueden reproducirse dentro de células huéspedes. El ejemplo de estas bacterias parece apoyar esta teoría, pues es probable que su dependencia del parasitismo haya causado la pérdida de los genes que les permitían sobrevivir fuera de una célula. También se le llama «teoría de la degeneración».[47] [48]
- Teoría del origen celular (también llamada «hipótesis del nomadeo» o «hipótesis del vagabundeo»):[47] [49] algunos virus podrían haber evolucionado a partir de fragmentos de ADN o ARN que «escaparon» de los genes de un organismo mayor. El ADN fugitivo podría haber provenido de plásmidos (fragmentos de ADN que pueden moverse entre células) o transposones. Estos son moléculas de ADN que se replican y se mueven a diferentes posiciones en el interior de los genes de la célula.[50] Antiguamente llamados «genes saltarines», son ejemplos de elementos móviles genéticos y podrían ser el origen de algunos virus. Los transposones fueron descubiertos en 1950 por Barbara McClintock a partir de sus estudios en maíz.[51]
- Teoría de la coevolución: los virus podrían haber evolucionado de complejas moléculas de proteínas y ácido nucleico, al mismo tiempo que aparecieron las primeras células en la Tierra, y habrían sido dependientes de la vida celular durante muchos millones de años. Los viroides son moléculas de ARN que no son clasificadas como virus porque carecen de envoltura proteica. Sin embargo, tienen características comunes a diversos virus y a menudo se les llama agentes subvíricos.[52] Los viroides son importantes patógenos de las plantas.[53] No codifican proteínas, pero interactúan con la célula huésped y la utilizan para replicarse y producir sus proteínas.[54] El virus de la hepatitis D de los humanos tiene un genoma de ARN similar al de los viroides pero tiene un envoltorio proteico derivado del virus de la hepatitis B y no puede producir uno propio. Por lo tanto, es un virus defectuoso que no puede replicarse sin la ayuda del virus de la hepatitis B.[55] De la misma forma, el virófago 'sputnik' es dependiente del mimivirus, el cual infecta a Acanthamoeba castellanii.[56] Estos virus que dependen de otras especies víricas reciben el nombre de satélites, y podrían representar estadios evolutivos intermedios entre los viroides y los virus.[57] [58]
Los priones son moléculas proteicas infecciosas que no contienen ni ADN ni ARN.[59] En las ovejas, causan una infección llamada tembladera ovina, y en el ganado vacuno causan encefalopatía espongiforme bovina (la «enfermedad de las vacas locas»). En los humanos, causan kuru y la enfermedad de Creutzfeldt-Jakob.[60] Son capaces de replicarse pues algunas proteínas pueden existir en dos formas diferentes y los priones cambian la forma normal de una proteína huésped en la forma del prion. Esto inicia una reacción en cadena en la que cada proteína priónica convierte muchas proteínas del huésped en más priones, y estos priones convierten a su vez aún más proteínas en priones. Aunque son fundamentalmente diferentes de los virus y los viroides, su descubrimiento da credibilidad a la teoría de que los virus podrían haber evolucionado de moléculas autoreplicadoras.[61]
El análisis informático de secuencias de ADN de los virus y los huéspedes está produciendo una mejor comprensión de las relaciones evolutivas entre diferentes virus y podría ayudar a identificar los antepasados de los virus modernos. En la actualidad, estos análisis no han ayudado a decidir cuál (o cuáles) de las teorías previamente esbozadas es correcta. Sin embargo, parece improbable que todos los virus actualmente conocidos compartan un antepasado común y probablemente los virus han aparecido en varias ocasiones en el pasado por medio de uno o más mecanismos, con lo cual, podrían ser correctas todas ellas.[62]
Microbiología
Propiedades de vida
Existen opiniones dispares sobre si los virus son una forma de vida o estructuras orgánicas que interactúan con los seres vivos. Por ello algunos autores se refieren a ellos como «organismos al límite de la vida».[63] Por una parte se asemejan a los organismos que tienen genes y evolucionan por selección natural,[64] y se reproducen creando múltiples copias de sí mismos para autoensamblarse. Sin embargo, carecen de estructura celular, lo cual es considerado la unidad básica de la vida. Además, los virus no tienen un metabolismo propio, y necesitan una célula hospedadora para crear nuevos productos. Por tanto, no se pueden reproducir en el exterior de una célula huésped (aunque bacterias como Rickettsia y Chlamydia son considerados organismos vivos a pesar de tener la misma limitación). Las formas de vida aceptadas utilizan la división celular para reproducirse, mientras que los virus aparecen de forma súbita y en gran cantidad dentro de las células, lo que es análogo al crecimiento autónomo de los cristales. El autoensamblaje de los virus dentro de las células tiene implicaciones para el estudio del origen de la vida, pues refuerza las hipótesis de que la vida podría haber comenzado en forma de moléculas orgánicas autoensamblantes.[65]
Ácido nucleico
El ácido nucleico en los virus contiene la información específica y el potencial para modificar operaciones en la célula infectada.[66] Los ácidos nucleicos son macromoléculas constituidas por cadenas de nucleótidos, los cuales a su vez están constituidos por una base nitrogenada asociada a un azúcar del grupo de las pentosas y a uno o más grupos de fosfatos. Existen cuatro posibles tipos de ácido nucleico viral:[67] ADN de cadena sencilla, ADN de cadena doble, ARN de cadena sencilla y ARN de cadena doble. Virus que contienen cualquiera de estos tipos de ácido nucleico pueden ser encontrados tanto entre los fagos como entre los virus que infectan a plantas o animales.[68]
El ADN de algunos bacteriófagos se caracteriza por contener bases raras que substituyen alguna o algunas de las bases normalmente presentes en el ADN.[68] El ADN de cadena doble presente en algunos virus, se caracteriza por tener segmentos de cadena sencilla en ambos extremos de la molécula. Debido a que son complementarias las secuencias de nucleótidos presentes en ambos extremos. El ADN naturalmente circular puede ser de cadena sencilla como en el fago ØXI74,[68] o de cadena doble, como en el virus SV4O. Existe evidencia de que algunos virus ARN que producen infecciones en vegetales como el limonero y la papa contienen moléculas circulares de ARN.
En los últimos diez años se han desarrollado una variedad de técnicas y métodos que permiten determinar la secuencia de nucleótidos en cualquier tipo de ácido nucleico. La primera secuencia completa de un ARN viral fue determinada en el fago MS2 por el grupo de Walter Fiers en 1976.[68] En 1977, Fred Sanger y colaboradores publicaron la secuencia completa del genoma del fago ØXl74,[68] constituido por ADN de cadena sencilla. Posteriormente, muchos otros genomas virales de mayor tamaño y complejidad han sido secuenciados en parte o en su totalidad.
Estructura
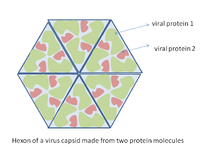 Diagrama de cómo se puede construir una cápside vírica a partir de múltiples copias de sólo dos moléculas proteicas.
Diagrama de cómo se puede construir una cápside vírica a partir de múltiples copias de sólo dos moléculas proteicas.Los virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.[69] La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.[70] Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.[71]
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros.[72] Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.[73] [74] Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.[75] [76] En general, hay cuatro tipos principales de morfología vírica:
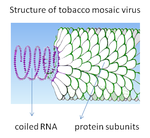 Diagrama de la estructura del virus del mosaico del tabaco: el ARN viral está enrollado en la hélice formada por subunidades proteicas repetidas.
Diagrama de la estructura del virus del mosaico del tabaco: el ARN viral está enrollado en la hélice formada por subunidades proteicas repetidas.- Helicoidal
- las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.[77]
- Icosaédrica
- la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones.[78]
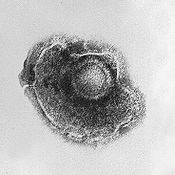 El virus del Herpes tiene una envoltura lípida.
El virus del Herpes tiene una envoltura lípida.- Envoltura
- algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar.[79]
- Complejos
- los virus tienen una cápside que no es ni puramente helicoidal, ni puramente icosaédrica, y que puede poseer estructuras adicionales como colas proteicas o una pared exterior compleja. Algunos bacteriófagos (como el Fago T4) tienen una estructura compleja que consiste en un cuerpo icosaédrico unido a una cola helicoidal (esta cola actúa como una jeringa molecular, atacando e inyectando el genoma del virus a la célula huésped),[80] que puede tener una base hexagonal con fibras caudales proteicas que sobresalgan.
Los poxvirus son virus grandes y complejos con una morfología inusual. El genoma vírico está asociado con proteínas dentro de una estructura discal central conocida como nucleoide. El nucleoide está rodeado por una membrana y dos cuerpos laterales de función desconocida. El virus tiene una envoltura exterior con una espesa capa de proteína en la superficie. La partícula en general es ligeramente pleomorfa, con una forma que puede ir de la de un huevo a la de un ladrillo.[81] Mimivirus es el virus más grande conocido, con un diámetro en su cápside de 400 nm. De su superficie se proyectan filamentos proteicos de 100 nm. La cápside, tomada en microscopio electrónico, tiene una forma hexagonal, de manera que probablemente es icosaédrica.[82]
Algunos virus que infectan a las Archaeas tienen estructuras inusuales, que no están relacionadas con ningún otro virus conocido. De igual manera, algunos bacteriófagos pueden tener diferentes estructuras en cuanto a su cola, con formas algo raras con respecto a otros virus.[83]
Genoma
Diversidad del genoma de los virus Propiedad Parámetros Ácido nucleico - ADN
- ARN
- Ambos AND y ARN (Tanto ADN como ARN)
Forma - Lineal
- Circular
- Segmentada
Cadenas - Monocatenarias
- Bicatenarias
- Bicatenarias con regiones monocatenarias
Sentido - Sentido positivo (+)
- Sentido negativo (−)
- Ambos sentidos (+/−)
Se puede ver una enorme variedad de estructuras genómicas entre las especies de virus que, como grupo, contienen una diversidad genómica superior a la de los reinos de plantas, los animales o las bacterias. Hay millones de diferentes tipos de virus;[3] y únicamente alrededor de 5.000 de ellos han sido descritos detalladamente.[2] Un virus tiene un genoma compuesto de ADN o bien de ARN, y reciben respectivamente los nombres de «virus ADN» y «virus ARN». La gran mayoría de virus utilizan el ARN. Los virus de las plantas tienden a tener ARN monocatenario y los bacteriófagos tienden a tener ADN bicatenario.[84]
Los genomas víricos pueden ser circulares, como los polyomaviridae o lineales, como los adenoviridae. El tipo de ácido nucleico es irrelevante para la forma del genoma. En los virus ARN, el genoma a menudo está dividido en partes separadas dentro del virión, y se le califica de «segmentado». Cada segmento suele codificar una proteína y los segmentos suelen estar reunidos en una cápside. No es necesario que cada segmento se encuentre en el mismo virión porque el virus en general es infeccioso, como lo demuestra el Brome mosaic virus.[69]
Poco importa el tipo de ácido nucleico, un genoma vírico puede ser o bien monocatenario o bicatenario. Los genomas monocatenarios consisten en un ácido nucleico no emparejado, similar a la mitad de una escalera de mano cortada por la mitad. Los virus bicatenarios consisten en dos ácidos nucleicos emparejados y complementarios, similares a una escalera de mano entera. Algunos virus, como los Hepadnaviridae, contienen un genoma que es parcialmente bicatenario y parcialmente monocatenario.[84]
En los virus ARN o los virus ADN monocatenarios, las cadenas pueden ser o bien positivas (cadenas plus) o negativas (cadenas minus), dependiendo de si son complementarias en el ARN mensajero (ARNm) vírico. El ARN viral positivo es idéntico al ARNm viral y por tanto puede ser traducido inmediatamente por la célula huésped. El ARN viral negativo es complementario del ARNm y por tanto debe ser convertido en ARN positivo por una ARN polimerasa antes de ser traducido. La nomenclatura del ADN es similar a la del ARN, en cuanto a la «cadena codificadora» del ARNm vírico que le es complementaria (-), y la «cadena no codificadora» que es una copia (+).[84]
El tamaño del genoma varía mucho entre especies. Los genomas víricos más pequeños sólo codifican cuatro proteínas y pesan unos 106 daltons; los más grandes pesan unos 108 daltons y codifican más de un centenar de proteínas.[84] Los virus ARN suelen tener genomas más pequeños que los virus ADN debido a una tasa de error más alta a la hora de replicarse, y tienen un límite superior de tamaño. Por encima de este límite, los errores en la replicación del genoma hacen que el virus sea inofensivo o incluso, incompetente. Para compensar esto, los virus ARN a menudo inician un proceso de segmentación en el que el genoma es separado en moléculas más pequeñas, reduciendo así las posibilidades de error. En cambio, los virus ADN tienen genomas mayores gracias a la elevada fidelidad de sus enzimas de replicación.[85]
Los virus sufren cambio genético por diversos mecanismos. Estos incluyen un proceso llamado deriva genética en el que las bases individuales del ADN o el ARN mutan en otras bases. La mayoría de estas mutaciones puntuales son imperceptibles pues la proteína que codifica el gen no cambia, pero aún así, puede conferir ventajas evolutivas como resistencia a los medicamentos antivíricos.[86] El cambio antigénico se produce cuando hay un cambio significativo en el genoma del virus. Esto ocurre como resultado de una recombinación genética. Cuando esto se produce en los virus de la gripe, pueden resultar pandemias.[87] Los virus ARN suelen existir como quasiespecies o en enjambres de virus de la misma especie pero con secuencias de nucleósidos del genoma ligeramente diferentes. Estos grupos son un objetivo destacado por la selección natural.[88]Los genomas segmentados ofrecen ventajas evolutivas; diferentes cepas de un virus con el genoma segmentado pueden intercambiar y combinar genes, produciendo virus progenénicos (o descendientes) con características únicas. Esto recibe el nombre de «sexo vírico».[89]
La recombinación genética es el proceso por el cual una cadena de ADN es rota y luego unida al extremo de una molécula de ADN diferente. Esto se puede producir cuando diferentes virus infectan las mismas células al mismo tiempo, y estudios de la evolución de los virus han demostrado que la recombinación tiene un papel muy importante en las especies estudiadas.[90] La recombinación es común en los virus ARN y ADN.[91] [92]
Ciclo de los virus
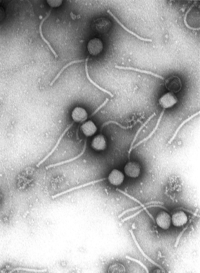
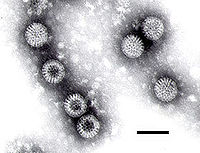 Micrografía electrónica de un rotavirus. La barra mide 100 nm.
Micrografía electrónica de un rotavirus. La barra mide 100 nm.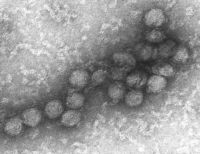
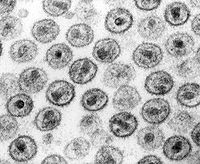 Micrografía del Virus de la inmunodeficiencia humana.
Micrografía del Virus de la inmunodeficiencia humana. Micrografía hecha del virus de Ébola.
Micrografía hecha del virus de Ébola.- La adhesión o adsorción es una unión específica entre proteínas de la cápside vírica y receptores específicos de la superficie celular del huésped, pero algunos bacteriófagos también son capaces de adherirse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera. Para que esto suceda la bacteria debe contener el factor sexual "F" o ciertas colicinas (factores de resistencia contra agentes antimicrobianos). Los bacteriófagos filamentosos con ADN de cadena sencilla se adhieren a las puntas de estos pili mientras que los bacteriófagos esféricos de ARN se adhieren a los costados de éstos. La especificidad de unión —proteína y cápside— se determina por la variedad de huéspedes de los virus. Por ejemplo, el VIH sólo infecta linfocitos T humanos, pues su proteína de superficie, gp120, puede interactuar con la CD4 y con receptores de la superficie del linfocito T. Este mecanismo ha evolucionado para favorecer los virus que sólo pueden infectar células en que se pueden replicar. La adhesión al receptor que puede inducir cambios en la proteína de la envoltura viral que resultan en la fusión de las membranas viral y celular.
- La penetración sigue a la adhesión;[n. 3] los virus se introducen en la célula huésped mediante endocitosis mediada por receptores o por fusión de membrana. Esto recibe a menudo el nombre de penetración vírica. La infección de las células vegetales es diferente a la de las células animales. Las plantas tienen una pared celular rígida hecha de celulosa y los virus sólo pueden entrar en las células cuando se produce un trauma en la pared celular.[93] Los virus como el virus del mosaico del tabaco también pueden moverse directamente a las plantas, entre células, a través de poros llamados plasmodesmos.[94] Las bacterias, como las de las plantas, tienen una fuerte pared celular que los virus tienen que romper para infectar la célula. Algunos virus han evolucionado mecanismos para inyectar su genoma a la célula bacteriana mientras la cápside viral permanece en el exterior.[95]
- El despojo es el proceso en que la cápside vírica es degradada por enzimas virales o del huésped, liberando así el ácido nucleico del genoma vírico.
- La replicación implica la síntesis de ARN mensajero (ARNm) vírico en todos los virus con rasgos de ARN positivos, la síntesis de proteínas víricas, el ensamblaje de proteínas víricas y la replicación del genoma viral. El proceso de replicación es esencial para mantener la estabilidad de la información genética contenida en el ADN. Esta replicación utiliza enzimas idénticas a las involucradas en la replicación del ADN celular y una característica común es la presencia de estructuras circulares temporales por lo menos en algunas de dicho proceso.
- Tras el ensamblaje de partículas víricas, a menudo se produce una modificación postraduccional de las proteínas víricas. En virus como el VIH, esta modificación (a veces llamada «maduración»), se produce después de que el virus haya sido liberado de la célula huésped.[96] El ensamblaje puede producir nuevas partículas virales. Los virus pueden autoensamblarse en un proceso similar a la cristalización, ya que las partículas virales, al igual que los cristales, constituyen estructuras que se encuentran en un estado mínimo de energía libre. Sin embargo, el genoma viral también puede especificar ciertos factores "morfogenéticos" que no contribuyen directamente a formar la estructura del virión, pero son necesarios para el proceso de ensamblaje. El fenómeno de autoensamblaje ocurre en la formación de diversas estructuras biológicas.
- Los virus son liberados de la célula huésped por lisis—un proceso que mata la célula reventando su membrana. Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Tipos de virus
El material genético y el método por el cual los virus se replican, varían entre los diferentes tipos.
- Virus ADN
- La replicación del genoma de la mayoría de virus ADN se produce en el núcleo de la célula. Si la célula tiene el receptor adecuado a la superficie, estos virus entran por fusión con la membrana celular o por endocitosis. La mayoría de virus ADN son completamente dependientes de la maquinaria de síntesis de ADN y ARN de la célula huésped, y su maquinaria de procesamiento de ARN. El genoma vírico debe atravesar la membrana nuclear de la célula para acceder a esta maquinaria.[97] [98] [99] [100]
-
-
- Virus ADN bicatenario
- Este tipo de virus tiene su material genético compuesto por ADN de doble cadena y se replica usando una ADN polimerasa,[98] que es dependiente del ADN y no del ARN. Este tipo de virus, por lo general, debe entrar en el núcleo de la célula huésped antes de que sea capaz de replicarse. Además, estos virus requieren de las polimerasas de la célula huésped para replicar el genoma viral y, por lo tanto, son altamente dependientes del ciclo celular.[99] Para que pueda realizarse la infección y la producción de progenie del virus se requiere que la célula esté en la fase de replicación, que es cuando las polimerasas de la célula están activas. El virus puede forzar a la célula a realizar la división celular y de forma crónica esto puede conducir a la transformación de la célula y, en última instancia, producir cáncer.[101] [102] [100]
-
-
-
- Virus ADN monocatenario
- Este tipo de virus posee en su material genético ADN de cadena sencilla y se replica usando una ADN polimerasa dependiente del ADN —al igual que el Virus ADN bicatenario.[102] A diferencias de los virus ADN bicatenarios, éstos poseen un ADN infectante monocatenario (de cadena simple), es decir, formado por una única cadena de nucleótidos, en lugar de la habitual doble hélice. Para que exista la replicación en este virus, es necesario que el ADN de cadena simple se convierta en ADN de cadena doble en las células infectadas.[99]
-
- Virus ARN
- Los virus ARN son únicos porque su información genética está codificada en ARN; esto quiere decir que usan el ácido ribonucleico (ARN) como material genético, o bien que en su proceso de replicación necesita el ARN. La replicación se suele producir en el citoplasma. Los virus ARN se pueden clasificar en unos cuatro grupos según su modo de replicación. La polaridad del ARN (si puede ser utilizado directamente o no para producir proteínas) determina en gran medida el mecanismo de replicación, y si el material genético es monocatenario o bicatenario. Los virus ARN utilizan sus propias ARN replicases para crear copias de su genoma.[103]
-
-
- Virus ARN bicatenario
- El virus ARN bicatenario es virus que posee ARN de cadena doble en su genoma. Como la mayoría de los virus ARN, se replican en el citoplasma y no dependen de las polimerasas de la células huésped como lo hacen los virus ADN, pues incluyen estas enzimas en el virión.[102] La traducción suele ser monocistrónica, lo que significa que cada uno de los segmentos codifica una sola proteína, a diferencia de otros virus que exhiben una traducción más compleja. Una características partícular de éstos es su capacidad para llevar a cabo la transcripción de los segmentos de ARN bicatenarios bajo las condiciones apropiadas dentro de la cápside.
-
-
-
- Virus ARN monocatenario positivo
- Los virus ARN monocatenarios positivos tienen ácido ribonucleico (ARN) de cadena sencilla de sentido positivo como material genético y no se replican usando ADN intermedio. Los virus ARN positivos son idénticos al ARNm viral y por lo tanto pueden ser inmediatamente traducidos por la célula huésped. Aunque el ARN purificado de un virus positivo puede causar directamente una infección, es menos infeccioso que el virus completo. La replicación tiene lugar principalmente en el citoplasma y no es tan dependiente del ciclo celular como en los virus ADN. Los virus ARN de sentido positivo tienen genomas con la misma polaridad del ARNm y pueden ser empleados directamente para la síntesis de proteínas usando la maquinaria de traducción de la célula huésped. Una de estas proteínas codificadas es la ARN replicasa, una ARN polimerasa que copia el ARN viral sin necesidad de pasar por una cadena de ADN intermedia.[102]
-
-
-
- Virus ARN monocatenario negativo
- Este virus tiene ácido ribonucleico (ARN) de cadena sencilla de sentido negativo como material genético y no se replica usando ADN intermedio.[102] El ARN viral negativo es complementario del ARNm y por lo tanto debe convertirse en ARN positivo por una ARN polimerasa antes de la traducción. El ARN purificado de un virus negativo no es por sí mismo infeccioso puesto que necesita ser traducido en ARN positivo. Los virus ARN de sentido negativo utilizan una ARN polimerasa o transcriptasa para formar ARN de sentido positivo. Esto significa que el virus debe aportar la enzima ARN polimerasa puesto que ésta es dependiente del ARN. La molécula ARN de sentido positivo entonces actúa como un ARNm viral, que se traduce en proteínas por los ribosomas del huésped. Las proteína resultante se dedica directamente a la producción de los elementos de los nuevos viriones, tales como las proteínas de la cápside y la ARN replicasa, que se encarga de la producción de nuevas moléculas de ARN de sentido negativo.
-
-
-
- Virus ARN monocatenario retrotranscrito
- Un virus ARN monocatenario retrotranscrito (o virus ssRNA-RT) es un virus con ARN de cadena sencilla en su genoma que se replica en la célula huésped mediante transcripción inversa, es decir, mediante la formación de ADN a partir del molde ARN.[102] Estos virus usan transcriptasa inversa codificada viralmente, es decir, una ADN polimerasa dependiente del ARN, para producir ADN a partir del genoma ARN viral. Este ADN a menudo se integra en el genoma del huésped, como en el caso de los retrovirus y seudovirus, donde es replicado y transcrito por el huésped.
-
-
-
- Virus ADN bicatenario retrotranscrito
- Los virus de transcripción inversa se replican mediante la transcripción inversa, que es la formación de ADN a partir de una plantilla de ARN.[102] Los virus de transcripción inversa que contienen un genoma de ARN utilizan un intermedio de ADN para replicarse, mientras que los que contienen un genoma de ADN utilizan un intermedio de ARN durante la replicación del genoma.
-
Efectos en la célula huésped
La variedad de efectos estructurales y bioquímicos de los virus sobre las células huésped es grande.[104] Reciben el nombre de «efectos citopáticos».[105] La mayoría de infecciones víricas acaban provocando la muerte de la célula huésped, entre cuyas causas están la lisis de la célula, las alteraciones de la membrana superficial de la célula y la apoptosis.[106] A menudo, la muerte de la célula es causada por el paro de sus actividades normales debido a la supresión por proteínas específicas del virus, que no son todas componentes de la partícula vírica.[107]
Algunos virus no causan cambios aparentes en la célula infectada. Las células en que los virus es latente e inactivo presentan pocos signos de infección y a menudo funcionan normalmente.[108] Esto causa infecciones persistentes y el virus a menudo permanece durmiente durante muchos meses o años. Este suele ser el caso del herpes simple.[109] [110] Algunos virus, como el virus de Epstein-Barr, a menudo hacen proliferar las células sin causar malignidad,[111] pero otros, como los papilomavirus, son una causa demostrada de cáncer.[112]
Clasificación
Las clasificaciones intentan describir la diversidad de virus dándoles nombre y agrupándolos según sus semejanzas. En 1962, André Lwoff, Robert Horne y Paul Tournier fueron los primeros en desarrollar una forma de clasificación de los virus, basada en el sistema jerárquico linneano.[113] Este sistema basa la clasificación en filos, clases, órdenes, familias, géneros y especies. Los virus fueron agrupados según sus propiedades compartidas (no las de sus huéspedes) y el tipo de ácido nucleico del que se compone su genoma.[114] Posteriormente se formó Comité Internacional de Taxonomía de Virus.
Clasificación del ICTV
El Comité Internacional de Taxonomía de Virus (ICTV) desarrolló el sistema de clasificación actual y escribió pautas que daban más importancia a ciertas propiedades de los virus para mantener la uniformidad familiar. Un sistema universal para clasificar los virus y una taxonomía unificada han sido establecidos desde 1966. El 7 º Informe del ICTV formalizó por primera vez el concepto de especie vírica como el taxón más bajo de una jerarquía ramificada de taxones de virus.[n. 4] [115] Sin embargo, actualmente sólo se ha estudiado una pequeña parte de toda la diversidad de los virus, y análisis de muestras obtenidas de humanos revelan que aproximadamente un 20% de secuencias víricas recuperadas no han sido observadas anteriormente. Muestras del ambiente, como sedimentos marinos y oceánicos, revelan que la gran mayoría de secuencias son completamente nuevas.[116]
La estructura general de la taxonomía es la siguiente:
La taxonomía actual del ICTV (2008) reconoce cinco órdenes: los caudovirales, los herpesvirales, los mononegavirales, los nidovirales y los picornavirales. El comité no distingue formalmente entre subespecies, cepas y aislamientos. En total, hay cinco órdenes, 82 familias, 11 subfamilias, 307 géneros, 2.083 especies y unos 3.000 tipos que aún no han sido clasificados.[117] [118]
Clasificación Baltimore
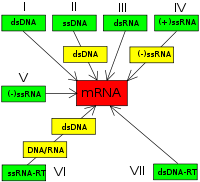 Esquema de la clasificación Baltimore de los virus.
Esquema de la clasificación Baltimore de los virus.David Baltimore, biólogo ganador del Premio Nobel, diseñó el sistema de clasificación que lleva su nombre.[37] [119] El sistema de clasificación del ICTV es utilizado en combinación con el sistema de clasificación de Baltimore en la clasificación moderna de los virus.[120] [121] [122]
La clasificación de Baltimore de los virus se basa en el mecanismo de producción de ARNm. Los virus deben generar ARNm de su genoma para producir proteínas y replicarse, pero cada familia de virus utiliza mecanismos diferentes. El genoma de los virus puede ser monocatenario (ss) o bicatenario (ds), de ARN o ADN, y pueden utilizar o no la transcriptasa inversa. Además, los virus ARN monocatenarios pueden ser o positivos (+) o negativos (-). Esta clasificación reparte los virus en siete grupos:
-
- I: Virus dsDNA (ej., adenovirus, herpesvirus, poxvirus)
-
- II: Virus ssDNA (ej., parvovirus)
-
- III: Virus dsARN (ej., reovirus)
-
- IV: Virus (+)ssRNA (ej., picornavirus, togavirus)
-
- V: Virus (-)ssRNA (ej., Ortomixovirus, rabdovirus)
-
- VI: Virus ssRNA-RT (ej., retrovirus)
-
- VII: Virus dsDNA-RT (ej., Hepadnaviridae)
Como ejemplo de la clasificación vírica, el virus de la varicela, varicela zoster (VZV), pertenece al orden de los herpesvirales, la familia de los Herpesviridae, la subfamilia de los Alphaherpesvirinae y el género Varicellovirus. El VZV se encuentra en el grupo I de la clasificación de Baltimore porque es un virus ADN bicatenario que no utiliza la transcriptasa inversa.
Virus y enfermedades humanas
Ejemplos de enfermedades humanas comunes provocadas por virus incluyen el resfriado, la gripe, la varicela y el herpes simple.[125] [126] [127] Muchas enfermedades graves como el ébola, el sida, la gripe aviar y el SARS son causadas por virus.[128] [129] [130] La capacidad relativa de los virus de provocar enfermedades se describe en términos de «virulencia». Otras enfermedades están siendo investigadas para descubrir si su agente causante también es un virus, como la posible conexión entre el herpesvirus humano 6 (HHV6) y enfermedades neurológicas como la esclerosis múltiple y el síndrome de fatiga crónica.[131] Actualmente existe un debate sobre si el bornaviridae, antiguamente considerado la causa de enfermedades neurológicas en los caballos, podría ser la causa de enfermedades psiquiátricas en los humanos.[132]
Los virus tienen diferentes mecanismos mediante los cuales causan enfermedades a un organismo, que dependen en gran medida en la especie de virus. Los mecanismos a nivel celular incluyen principalmente la lisis de la célula, es decir, la ruptura y posterior muerte de la célula. En los organismos pluricelulares, si mueren demasiadas células del organismo en general comenzará a sufrir sus efectos. Aunque los virus causan una disrupción de la homeostasis saludable, provocando una enfermedad, también pueden existir de manera relativamente inofensiva en un organismo. Un ejemplo sería la capacidad del virus del herpes simple de permanecer en un estado durmiente dentro del cuerpo humano. Esto recibe el nombre de «latencia»[133] y es una característica de todos los herpesvirus, incluyendo el virus de Epstein-Barr (que causa mononucleosis infecciosa) y el virus de la varicela zoster (que causa la varicela). Las infecciones latentes de varicela pueden generarse posteriormente en la etapa adulta del ser humano en forma de la enfermedad llamada herpes zóster.[134] Sin embargo, estos virus latentes algunas veces suelen ser beneficiosos, incrementando la inmunidad del cuerpo contra algunos seres patógenos, como es el caso del Yersinia pestis.[135] Cuando alguna enfermedad viral vuelve a reincidir en cualquier etapa de la vida se conoce popularmente como culebrilla.
Algunos virus pueden causar infecciones permanentes o crónicas, en que los virus continúan replicándose en el cuerpo a pesar de los mecanismos de defensa del huésped.[136] Esto es habitual en las infecciones de virus de la hepatitis B y de la hepatitis C. Los enfermos crónicos son conocidos como portadores, pues sirven de reservorio de los virus infecciosos.[137] En poblaciones con una proporción elevada de portadores, se dice que la enfermedad es endémica.[138] Algunos virus pueden mutar dentro de las células huéspedes, reforzando sus defensas contra diversos antivirales, proceso conocido como mutación.[139]
Epidemiología
La epidemiología viral es la rama de la ciencia médica que estudia la transmisión y el control de infecciones víricas en los humanos. La transmisión de virus puede ser vertical (de madre a hijo) u horizontal (de una persona a otra). Ejemplos de transmisión vertical incluyen el virus de la hepatitis B o el VIH, en que el bebé ya nace infectado con el virus.[140] Otro ejemplo más raro es el virus de la varicela zóster. Normalmente causa infecciones relativamente leves en los humanos, pero puede resultar fatal para los fetos y los bebés recién nacidos.[141] La transmisión horizontal es el mecanismo de contagio de virus más extendido. La transmisión puede ser por intercambio de sangre o por el cambio de fluidos en la actividad sexual (ej., VIH, hepatitis B y hepatitis C), por la boca por el intercambio de saliva (ej., virus de Epstein-Barr), por alimentos o agua contaminados (ej., norovirus), por la respiración de virus en forma de aerosol (ej., virus de la gripe) o por insectos vectores como los mosquitos (ej., dengue o malaria). La tasa y la velocidad de la transmisión de infecciones víricas dependen de factores como la densidad de población, el número de individuos susceptibles (los que no son inmunes),[142] la calidad del sistema sanitario y el tiempo.[143]
La epidemiología se utiliza para romper la cadena de infecciones en poblaciones durante brotes de enfermedades víricas.[144] Se utilizan medidas de control basándose en el conocimiento del modo de transmisión del virus. Una vez identificado el virus, a veces se puede romper la cadena de infecciones por medio de vacunas. Cuando no se puede contar con vacunas, pueden resultar eficientes el saneamiento y la desinfección. A menudo se aíslan las personas infectadas del resto de la comunidad, y los que han estado expuestos al virus son puestos en cuarentena.[145] Para controlar el brote de fiebre aftosa en bovinos británicos en 2001, se sacrificaron miles de cabezas de ganado.[146] La mayoría de infecciones víricas de los humanos y otros animales tienen un periodo de incubación durante el cual la infección no causa ningún signo o síntoma.[147] Los períodos de incubación de las enfermedades víricas van desde unos varios días hasta semanas, pero son conocidos en el caso de muchas infecciones.[148] Tras el periodo de incubación hay un «periodo de comunicabilidad», un tiempo durante el cual el individuo o animal infectado es contagioso y puede infectar otra persona o animal.[149] Este periodo también es conocido en muchas infecciones, y el conocimiento de la longitud de ambos periodos es importante en el control de brotes.[150] Cuando un brote causa una proporción inusualmente elevada de infecciones en una población, comunidad o región, se le llama epidemia. Si un brote se extiende en todo el mundo se le llama pandemia.[151]
Epidemias y pandemias
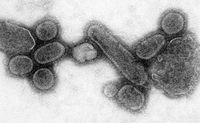 Reconstrucción del tenebroso virus de la gripe española que llegó a matar a cerca del 5% de la población humana entre 1918 y 1919. Es considerada la más grave pandemia en toda la historia de la humanidad.
Reconstrucción del tenebroso virus de la gripe española que llegó a matar a cerca del 5% de la población humana entre 1918 y 1919. Es considerada la más grave pandemia en toda la historia de la humanidad.Las poblaciones amerindias fueron devastadas por enfermedades contagiosas, especialmente la viruela, llevada a América por los colonos europeos. Es incierto el número de nativos americanos muertos por enfermedades extranjeras después de la llegada de Colón a América, pero se ha estimado que fue el 70% de la población indígena. Los estragos causados por esta enfermedad contribuyeron significativamente a los intentos de los europeos de ahuyentar o conquistar la población nativa.[152] [153] [154] [155] [156] [157] [158] Una pandemia es una epidemia global. La pandemia de gripe de 1918, a menudo llamada gripe española, fue una pandemia de gripe de categoría 5 provocada por un virus de la gripe A inusualmente grave y mortal. Las víctimas a menudo eran adultos jóvenes sanos, en contraste con la mayoría de brotes de gripe, que afectan predominantemente pacientes jóvenes, ancianos o débiles.[159] La pandemia de gripe española duró de 1918 a 1919. Las estimaciones más antiguas dicen que mató entre 40 y 50 millones de personas,[160] mientras que las más recientes sugieren que podría haber muerto hasta 100 millones de personas, o un 5% de la población mundial en 1918.[161]
La mayoría de investigadores creen que el VIH se originó en el África subsahariana durante el siglo XX;[162] y actualmente es una pandemia, con un número estimado de 38,6 millones de enfermos en todo el mundo.[163] El Programa Conjunto de las Naciones Unidas sobre el VIH/SIDA (UNAIDS) y la Organización Mundial de la Salud (OMS) estiman que el sida ha matado a más de 25 millones de personas desde que fue reconocida por primera vez el 5 de junio de 1981, siendo una de las epidemias más destructivas de la historia.[164] En 2007 hubo 2,7 millones de infecciones con VIH y dos muertes relacionadas con este virus.[165]
Algunos patógenos víricos muy letales son miembros de la familia de los Filoviridae. Los Filovirus son virus similares a filamentos que causan la fiebre hemorrágica vírica, e incluyen el Ébola y los virus de Marburg. El virus de Marburg atrajo la atención de la prensa en abril de 2005 por un brote en Angola. El brote, que comenzó en 2004 y se extendió en 2005, fue la peor epidemia del mundo de cualquier tipo de fiebre hemorrágica vírica.[166]
En 2009, surgió en México, una supuesta pandemia de Influenzavirus A (H1N1);[167] conocido como Virus H1N1/09 Pandémico.[168] El origen de la infección es una variante de la cepa H1N1, con material genético proveniente de una cepa aviaria, dos cepas porcinas y una humana que sufrió una mutación y dio un salto entre especies (o heterocontagio) de los cerdos a los humanos, y contagiándose de persona a persona.[169] La pandemia fue clasificada, según la OMS, de Nivel 6. Aproximadamente, murieron 14.286 en todo el mundo a causa de esta enfermedad.[170]
Cáncer
Los virus son una causa establecida de cáncer en los humanos y otras especies. Los cánceres virales son demasiado raros y sólo ocurren de unas cuantas personas (o animales). Los virus que producen cáncer pueden provenir de muchas familias, tanto de virus ADN como de virus ARN, y no únicamente del oncovirus (un término obsoleto para referirse a los retrovirus). El desarrollo del cáncer puede deberse a gran cantidad de factores como la debilidad inmunitaria del huésped y mutaciones en éste.[171] [172] Los virus más importantes asociados con cánceres humanos son el papilomavirus humano, el virus de la hepatitis B, el virus de Epstein-Barr, y el virus T-linfotrópico humano. El más reciente descubrimiento de un virus que causa cáncer es el poliomavirus (Merkel cell polyomavirus) que es la causa de un raro cáncer de piel denominado carcinoma de células de Merkel.[173]
Los virus de la hepatitis pueden causar una infección crónica que provoca cáncer de hígado.[174] [175] La infección con virus T-linfotrópico humano puede causar paraparesia espástica tropical y leucemia de linfocitos T del adulto.[176] Los papilomavirus humanos son una causa establecida de cáncer de cérvix, piel, ano y pene.[177] Dentro de los Herpesviridae, el human herpesvirus 8 causa sarcoma de Kaposi y linfoma de las cavidades corporales, y el virus de Epstein-Barr causa linfoma de Burkitt, enfermedad de Hodgkin, trastorno linfoproliferativo de los linfocitos B y carcinoma nasofaríngeo.[178] El Merkel cell poliomavirus está estrechamente relacionado con el SV40 y con los poliomavirus del ratón que han sido usados como modelos de animales para los virus del cáncer desde hace 50 años.[179]
Respuesta inmune del huésped
La primera línea de defensa del organismo contra los virus es el sistema inmunitario innato. Éste incluye las células y otros mecanismos que defienden al organismo de la infección de una forma no específica. Esto significa que las células del sistema innato reconocen y responden a los agentes patógenos de una manera genérica, pero, a diferencia del sistema inmune adaptativo, no confieren protección de larga duración o inmunidad.[180]
El ARN interferente es una importante defensa innata contra los virus.[181] Muchos virus tienen una estrategia de replicación que implica ARN bicatenario (dsRNA). Cuando tales virus infectan a una célula y liberan su molécula o moléculas de ARN, inmediatamente una proteína compleja denominada dicer se une al ARN y lo corta en pedazos más pequeños. Una vía bioquímica denominada complejo RISC se activa y degrada el ARNm viral. Los rotavirus evitan este mecanismo no desnudándose completamente dentro de la célula. El dsRNA genómico continúa protegido en el interior del núcleo del virión y se liberan los nuevos ARNm producidos a través de los poros de la cápside.[182] [183]
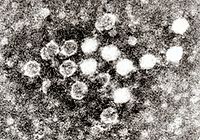
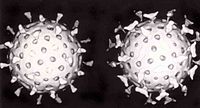 Dos rotavirus; el derecho está cubierto por anticuerpos que impiden la adhesión a la célula huésped.
Dos rotavirus; el derecho está cubierto por anticuerpos que impiden la adhesión a la célula huésped.Cuando el sistema inmunitario adaptativo de un vertebrado encuentra un virus, produce anticuerpos específicos que se unen al virus y lo hacen no infeccioso, lo que se denomina inmunidad humoral. Dos tipos de anticuerpos son importantes. El primero se denomina IgM y es altamente eficaz para neutralizar los virus, pero sólo es producido por las células del sistema inmune durante unas pocas semanas. El segundo, denominado IgG, se produce indefinidamente. La presencia de IgM en la sangre del huésped se utiliza para determinar una infección aguda, mientras que el IgG indica una infección en el pasado.[184] Los dos tipos de anticuerpos se analizan cuando se llevan a cabo las pruebas de inmunidad.[185]
Una segunda línea de defensa de los vertebrados frente a los virus se denomina inmunidad celular y consiste en las células inmunitarias conocidas como linfocitos T. Las células del organismo constantemente muestran cortos fragmentos de sus proteínas en la superficie celular. Si un linfocito T reconoce en una célula un fragmento sospechoso de ser viral, destruye dicha célula y a continuación se produce una proliferación de los linfocitos T específicos para ese virus. Los macrófagos son las células especialistas en la presentación antigénica.[186] [187] La producción de interferón es un importante mecanismo que interviene también en la defensa.[188]
No todas las infecciones por virus producen de esta manera una respuesta inmune protectora. El VIH evade al sistema inmunológico por el cambio constante de la secuencia de aminoácidos de las proteínas en la superficie del virión. Estos persistentes virus eluden el control mediante el secuestro y bloqueo de la presentación antigénica, resistencia a las citoquinas, evasión a las actividades de los lifocitos T, inactivación de la apoptosis, y el cambio antigénico.[189] Otros virus, denominados "virus neurotróficos", se propagan en el sistema neural, donde el sistema inmunológico puede ser incapaz de llegar a ellos.
Prevención
Dado que los virus utilizan la maquinaria de una célula huésped para reproducirse y residen en el interior, son difíciles de eliminar sin matar la célula huésped. Los enfoques médicos más eficientes para enfrentarse a las enfermedades víricas conocidos hasta ahora son las vacunas, que ofrecen resistencia a la infección, y los antivirales.
Vacunas
La vacunación es una forma barata y eficaz para la prevención de las infecciones causadas por los virus. Las vacunas se han utilizado para prevenir las enfermedades virales desde mucho antes al descubrimiento de los virus. Su uso ha dado lugar a una dramática disminución de la morbilidad (enfermedad) y mortalidad (muerte) asociada a infecciones virales como poliomielitis, sarampión, paperas y rubéola.[190] La viruela ha sido erradicada.[191] En la actualidad se dispone de vacunas para prevenir más de trece infecciones virales en los seres humanos,[192] y algunas más se utilizan para prevenir infecciones virales en animales.[193] El proceso de vacunación se basa en la idea de que se puede lograr inmunidad específica contra una enfermedad,[194] en particular si se provoca ésta en condiciones controladas de manera que el individuo no padece los síntomas asociados con dicha enfermedad y el sistema inmune reacciona produciendo un arsenal de anticuerpos y células inmunes con capacidad para destruir o neutralizar cualquiera otra invasión por parte del mismo agente infeccioso, como los linfocitos T; que son los responsables de coordinar la respuesta inmune celular.[195]
Las vacunas pueden consistir en virus vivos atenuados o en virus muertos, o en sólo las proteínas virales (antígenos).[196] Las vacunas vivas contienen formas debilitadas del virus que causa la enfermedad. Las vacunas vivas pueden ser peligrosas cuando se administran a las personas inmunodeficientes, puesto que en estas personas incluso el virus debilitado puede causar la enfermedad original.[197] Sin embargo, la vacuna contra el virus de la fiebre amarilla, obtenida de una cepa atenuada denominada 17D, es posiblemente una de las vacunas más seguras y eficaces fabricadas.
La biotecnología y las técnicas de ingeniería genética se utilizan para producir vacunas de subunidades. Estas vacunas usan sólo la cápside de proteínas del virus. La vacuna de la hepatitis B es un ejemplo de este tipo de vacuna.[198] Las vacunas de subunidades son seguras para pacientes inmunodeficientes, ya que no pueden causar la enfermedad.[199]
Medicamentos antivirales
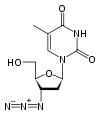 El fármaco antiviral zidovudina (AZT), análogo de la timidina.
El fármaco antiviral zidovudina (AZT), análogo de la timidina.El primer fármaco que se presentó como agente antiviral verdaderamente selectivo y con éxito fue el aciclovir;[200] que fue utilizado como tratamiento profiláctico del herpes genital y cutáneo, y también en el tratamiento de las lesiones causadas por el Herpes zoster.[201] Durante los últimos veinte años, el desarrollo de fármacos antivirales continuó aumentado rápidamente, impulsado por la epidemia del sida. Los medicamentos antivirales son a menudo «análogos de nucleósidos» (falsos nucleósidos, los bloques de construcción de los ácidos nucleicos) que los virus incorporan a sus genomas durante la replicación. El ciclo de vida del virus entonces se detiene debido a que las nuevas cadenas de ADN sintetizadas son defectuosas. Esto se debe a que los análogos carecen de los grupos hidroxilos que junto a los átomos de fósforo forman los enlaces de la fuerte «columna vertebral» de la molécula de ADN. A esto se le denomina interrupción de la cadena de ADN.[202] Ejemplos de análogos de nucleósidos son el aciclovir para tratar el virus del herpes y lamivudina para las infecciones de VIH y hepatitis B. Aciclovir es uno de los fármacos antivirales más antiguos y frecuentemente prescritos.[203]
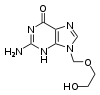 El fármaco antiviral aciclovir, análogo de la guanosina.
El fármaco antiviral aciclovir, análogo de la guanosina.La hepatitis C es causada por un virus ARN. En el 80% de las personas infectadas, la enfermedad es crónica y sin tratamiento continúan siendo infecciosas para el resto de sus vidas. Sin embargo, ahora existe un tratamiento efectivo con el fármaco ribavirina, un análogo de nucleósido, en combinación con interferón.[204] Actualmente se está desarrollando una estrategia similar con lamivudina para el tratamiento de los portadores crónicos de hepatitis B.[205] Otros fármacos antivirales en uso tienen como objetivo diferentes etapas del ciclo replicativo viral. El VIH depende de una enzima proteolítica denominada proteasa VIH-1 para ser plenamente infeccioso. Existe una clase de medicamentos denominados inhibidores de la proteasa que han sido diseñados para inactivar esta enzima.
El sida, provocado por el VIH, tiene un tratamiento antiviral de zidovudina (azidotimidina o AZT). La zidovudina es un potente inhibidor de la transcriptasa inversa (RT), enzima esencial en el proceso de replicación del VIH. Sin embargo, sus efectos no son duraderos y en algunos casos, éstos son inútiles, puesto que el VIH es un retrovirus y su genoma de ARN debe ser transcrito por la RT para convertirlo en una molécula de ADN que constituye el provirus. La zidovudina no tiene ningún efecto sobre el provirus, ya que sólo inhibe su formación más no la expresión de ésta en las células huéspedes. Por otra parte, el uso duradero de zidovudina podría provocar una mutación del VIH, haciendo resistente al virus a este tratamiento.
Infección en otras especies
Los virus infectan todo tipo de vida celular y, aunque los virus existen en todo el mundo, cada especie celular tiene un grupo de virus específico, que a menudo sólo infectan esta especie.[206] Los virus son importantes patógenos del ganado. Enfermedades como la fiebre aftosa y la lengua azul son causadas por virus.[207] Los animales de compañía (como perros, gatos y caballos), si no se les vacuna, son susceptibles a infecciones víricas graves. El parvovirus canino es causado por un pequeño virus ADN y las infecciones a menudo son fatales en los cachorros.[208] Como todos los invertebrados, la abeja de la miel es susceptible a muchas infecciones víricas.[209] Afortunadamente, la mayoría de virus coexisten de manera inofensiva con su huésped y no causan signos o síntomas de enfermedad.[210]
Plantas
Hay muchos tipos de virus de las plantas, pero a menudo sólo causan una pérdida de producción, y no es económicamente viable intentar controlarlos. Los virus de las plantas a menudo son transmitidos de una planta a otra por organismos conocidos como vectores. Normalmente son insectos, pero también se ha demostrado que algunos hongos, nemátodos y organismos unicelulares son vectores. Cuando se considera económico el control de infecciones por fitovirus (en los frutos perennes, por ejemplo), los esfuerzos se concentran en matar a los vectores y eliminar huéspedes alternativos como malas hierbas.[211] Los fitovirus son inofensivos para los humanos y demás animales, pues sólo se pueden reproducir en células vegetales vivas.[212]
Las plantas tienen mecanismos de defensa elaborados y eficientes contra los virus. Uno de los más eficientes es la presencia de los llamados genes de resistencia (R). Cada gen R confiere resistencia a un virus determinado desencadenando zonas localizadas de muerte celular alrededor de la célula infectada, que se pueden ver a simple vista en forma de manchas grandes. Esto detiene la expansión de la infección.[213] La interferencia del ARN también es una defensa efectiva en las plantas. Cuando están infectadas, las plantas a menudo producen desinfectantes naturales que matan los virus, como el ácido salicílico, el óxido nítrico y moléculas reactivas de oxígeno.[214]
Bacterias
Los bacteriófagos son un grupo extremadamente común y diverso de virus. Por ejemplo, los bacteriófagos son la forma más común de entidad biológica en los medios acuáticos; en los océanos hay hasta diez veces más de estos virus que de bacterias,[215] alcanzando niveles de 250 millones de bacteriófagos por milímetro cúbico de agua marina.[216] Estos virus infectan bacterias específicas uniéndose a moléculas receptoras de superficie y entrando en la célula. En un periodo corto de tiempo (en algunos casos en unos minutos), las polimerasas bacterianas empiezan a traducir ARN vírico en proteína. Estas proteínas se convierten en nuevos viriones dentro de la célula, proteínas colaboradoras que ayudan a parecerse nuevos viriones, o proteínas implicadas en la lisis celular. Los enzimas víricos colaboran en la destrucción de la membrana celular y, en el caso del fago T4, un poco más de veinte minutos después de la inyección ya se pueden liberar más de 300 fagos.[217]
El mecanismo principal por el que las bacterias se defienden de los bacteriófagos es produciendo enzimas que destruyen el ADN ajeno. Estas enzimas, llamadas endonucleases de restricción, fragmentan el ADN vírico que los bacteriófagos introducen en las células bacterianas.[218] Las bacterias también tienen un sistema que utiliza secuencias CRISPR para retener fragmentos del genoma de virus que se han encontrado en el pasado, lo que les permite impedir la replicación del virus mediante una forma de interferencia del ARN.[219] [220] Este sistema genético proporciona a las bacterias una inmunidad adquirida a las infecciones.
Archaea
Algunos virus se replican dentro de los arqueas, se trata de virus ADN que parecen no tener relación con ninguna otra forma de virus y que tienen una variedad de formas inusuales, como botellas, barras con un gancho o incluso lágrimas. Estos virus han sido estudiados en mayor detalle en los termófilos, en particular los órdenes Sulfolobales y Thermoproteales.[4] [83] La defensa contra estos virus pueden incluir la interferencia del ARN de secuencias de secuencias repetidas de ADN del genoma arqueobacterial que están relacionadas con los genes de los virus.[221] [222]
Aplicaciones
Ciencias de la vida y medicina
Los virus son importantes para el estudio de la biología molecular y celular, pues son sistemas sencillos que se pueden utilizar para manipular e investigar el funcionamiento de las células.[223] El estudio y el uso de los virus ha ofrecido información valiosa sobre aspectos de la biología celular.[224] Por ejemplo, los virus han resultado útiles en el estudio de la genética y han contribuido a comprender los mecanismos básicos de la genética molecular, como la replicación del ADN, la transcripción, la maduración del ARN, la traducción, el transporte de proteínas y la inmunología.
Los genetistas a menudo utilizan virus como vectores para introducir genes en células que están estudiando. Esto es útil para hacer que la célula produzca una sustancia ajena, o para estudiar el efecto de la introducción de un nuevo gen en el genoma. A este proceso se le denomina transducción. De manera similar, la viroterapia utiliza virus como vectores para tratar diversas enfermedades, pues pueden dirigirse específicamente a células y al ADN. Tiene un uso prometedor en el tratamiento del cáncer y en la terapia génica. Científicos del este de Europa han utilizado la terapia fágica como alternativa a los antibióticos desde hace un tiempo, y el interés por este enfoque está creciendo debido al alto nivel de resistencia a los antibióticos observado actualmente en algunas bacterias patógenas.[225]
Materiales científicos y nanotecnología
Las tendencias actuales en nanotecnología prometen hacer un uso mucho más versátil de los virus. Desde el punto de vista de un científico de materiales, los virus pueden ser considerados nanopartículas orgánicas. Su superficie porta herramientas específicas diseñadas para cruzar las barreras de la célula huésped. El tamaño y la forma de los virus, así como el número y la naturaleza de los grupos funcionales de su superficie, están definidos con precisión. Por tanto, los virus son utilizados habitualmente en ciencia de materiales como carcasas de modificaciones de superficie unidas de forma covalente. Una cualidad particular de los virus es que pueden ser diseñados por evolución dirigida. Las técnicas potentes desarrolladas por las ciencias de la vida están siendo la base de enfoques de ingeniería hacia los nanomateriales, abriendo una gran variedad de usos mucho más allá de la biología y la medicina.[226]
Debido a su tamaño, forma y estructuras químicas bien definidas, los virus han sido utilizados como moldes para organizar materiales a nanoescala. Ejemplos recientes incluyen el trabajo hecho en el Naval Research Laboratory de Washington D. C., utilizando partículas del Cowpea mosaic virus (CPMV) para amplificar señales en sensores basados en chips de ADN. En este uso, las partículas víricas separan las tinciones fluorescentes utilizadas con el fin de evitar la formación de dímeros no fluorescentes que actúen como extintores.[227] Otro ejemplo es el uso del CPMV como nanoplaca de pruebas para moléculas electrónicas.[228]
Armas
La capacidad de los virus de causar epidemias devastadoras en las sociedades humanas ha levantado la preocupación que se puedan convertir en armas biológicas. La preocupación aumentó después de que se consiguiera recrear el infame virus de la gripe española en un laboratorio.[229] El virus de la viruela devastó numerosas sociedades a lo largo de la historia antes de ser erradicado. Actualmente sólo existe en varios laboratorios seguros en diversos lugares del mundo,[230] pero los temores que pueda ser utilizado como arma no están totalmente infundados;[230] la vacuna de la viruela no es segura —durante los años anteriores a la erradicación de la viruela, cayó más gente gravemente enferma como resultado de la vacunación que por la propia viruela[231] — y la vacunación para la viruela ya no se practica.[232] Por este motivo, gran parte de la población humana actual casi no tiene resistencia a la viruela.[230]
Notas
- ↑ Renato Dulbecco, en 1975, definió a un virus como:
Un virus es un parásito intracelular obligatorio que puede ser considerado como un bloque de material genético (ya sea ADN o ARN) capaz de replicarse en forma autónoma, y que está rodeado por una cubierta de proteína y en ocasiones también por una envoltura membranosa que lo protege del medio y sirve como vehículo para la transmisión del virus de una célula a otra.Mientras que la pareja investigadora Luna y Darnell, en 1967, argumentaron su propia definición:Los virus son entidades cuyos genomas son elementos de ácido nucleico que se replican dentro de las células vivas utilizando para este fin la maquinaria sintética de la propia célula hospedera y provocando la síntesis de elementos especializados que pueden transferir el genoma viral hacia otras células.
Citado en:- Aranda-Anzaldo, A., D. Viza y R. G. Busnel, "Chemical inactivation of human immunodeficiency virus (HIV) in vitro", Journal of Virological Methods 37, pp. 71-82.
- Cann, A. J., y J. Karn, "Molecular Biology of HIV: new insights into the virus life-cycle." AIDS3 (suppl. 1), 1989, pp. s19-s34.
- ↑ En el virus del mosaico del tabaco (VMT), una proteína compuesta por 158 aminoácidos constituye la subunidad básica a partir de la cual se construye la cápside del virus. En dicha proteína cuando menos la mitad de los aminoácidos presentes en el interior de la macromolécula son de tipo hidrofóbico, mientras que en la superficie de la misma hay tan sólo cuatro grupos hidrofóbicos en un segmento constituido por 24 residuos de aminoácidos.
- ↑ El caso mejor estudiado de la penetración de un virus en la célula hospedera está representado por el caso del fago T2. La cola de este fago es contráctil y en su forma extendida consiste de 24 anillos de subunidades que forman una funda que rodea a un elemento central. Cada anillo consta de 6 subunidades pequeñas y 6 subunidades mayores. Después de la adsorción del fago a la pared celular, ocurre una contracción de la cola que resulta en una fusión de las subunidades pequeñas y grandes para dar 12 anillos de 12 subunidades. El núcleo de la cola no es contráctil, razón por la cual, es expulsado e impulsado a través de las capas externas de la bacteria; generando así que la cabeza del fago se contraiga y esto resulta en la inyección del ADN viral en la célula bacteriana. Este proceso posiblemente es facilitado por la presencia de la enzima lisozima en la cola del fago; enzima que es capaz de digerir las proteínas de las cubiertas bacterianas; además de eso, hay 144 moléculas de adenosina trifosfato (ATP) en la funda de la cola del fago; la energía para la contracción de esta funda proviene de la conversión de la ATP en adenosina difosfato (ADP) por medio de una reacción hidrolítica que libera un grupo fosfato de la ATP.
- ↑ Según la definición expresada en Fields Virology, "una especie de viral es una clase politética de virus que constituyen un linaje replicante y ocupan un nicho ecológico determinado". Una clase "politética" es aquella en que los miembros tienen diversas propiedades comunes, aunque no comparten necesariamente una única característica definidora. Los miembros de una especie de virus son definidos colectivamente por un grupo consensuado de propiedades. Así pues, las especies de virus difieren los taxones víricos más altos, que son clases "universales" y como tales están definidas por propiedades que son necesarias para ser miembro.
Referencias
- ↑ a b Dimmock p. 4
- ↑ a b Dimmock p. 49
- ↑ a b Breitbart M, Rohwer F (Junio de 2005). "Here a virus, there a virus, everywhere the same virus?". Trends Microbiol 13 (6): 278–84. doi:10.1016/j.tim.2005.04.003. PMID 15936660
- ↑ a b c Lawrence CM, Menon S, Eilers BJ, et al. (Mayo de 2009). "Structural and functional studies of archaeal viruses". J. Biol. Chem. 284 (19): 12599–603. doi:10.1074/jbc.R800078200. PMID 19158076. PMC 2675988
- ↑ Edwards RA, Rohwer F (Junio 2005). «Viral metagenomics». Nat. Rev. Microbiol. 3 (6): pp. 504–10. doi:. PMID 15886693.
- ↑ Villarreal, L. P. (2005) Viruses and the Evolution of Life. ASM Press, Washington DC ISBN 1-55581-309-7
- ↑ Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. (2007). Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series). Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 0-7817-8215-5. pág. 315
- ↑ Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. (2007). Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series). Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 0-7817-8215-5. pág. 47
- ↑ Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H (Agosto 2003). «Phage as agents of lateral gene transfer». Curr. Opin. Microbiol. 6 (4): pp. 417–24. doi:. PMID 12941415.
- ↑ Shors pp. 49–50
- ↑ Sepkowitz KA (Junio 2001). «AIDS--the first 20 years». N. Engl. J. Med. 344 (23): pp. 1764–72. doi:. PMID 11396444.
- ↑ Weiss RA (May 1993). «How does HIV cause AIDS?». Science (journal) 260 (5112): pp. 1273–9. PMID 8493571.
- ↑ Cecil, Russell (1988). Textbook of Medicine. Philadelphia: Saunders. pp. 1523, 1799. ISBN 0721618480.
- ↑ «Medmicro Chapter 52». Consultado el 21-02-2009.
- ↑ a b The Online Etymology Dictionary. «Virus». Consultado el 12 de enero de 2010.
- ↑ The Online Etymology Dictionary. «virulent, a.». Consultado el 12 de enero de 2010.
- ↑ The Online Etymology Dictionary. «viral.». Consultado el 12 de enero de 2010.
- ↑ "The History of the Germ Theory," The British Medical Journal vol. 1 no. 1415 (1888), p. 312.
- ↑ Madigan M, Martinko J (editors). (2005). Brock Biology of Microorganisms (11th ed. ed.). Prentice Hall. ISBN 0-13-144329-1.
- ↑ Shors pp. 76–77
- ↑ a b c d Collier p. 3
- ↑ Dimmock p.4–5
- ↑ Shors p. 589
- ↑ D'Herelle F (Septiembre de 2007). "On an invisible microbe antagonistic toward dysenteric bacilli": brief note by Mr. F. D'Herelle, presented by Mr. Roux. 1917. Res. Microbiol. 158(7):553–4. Epub 28 de julio de 2007. PMID 17855060
- ↑ Steinhardt, E; Israeli, C; and Lambert, R.A. (1913) "Studies on the cultivation of the virus of vaccinia" J. Inf Dis. 13, 294–300
- ↑ Collier p. 4
- ↑ Goodpasture EW, Woodruff AM, Buddingh GJ (1931). "The cultivation of vaccine and other viruses in the chorioallantoic membrane of chick embryos". Science 74, pp. 371–372 PMID 17810781
- ↑ Rosen FS (2004). "Isolation of poliovirus—John Enders and the Nobel Prize". New England Journal of Medicine, 351, pp. 1481–83 PMID 15470207
- ↑ From Nobel Lectures, Physics 1981–1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
- ↑ Buist JB Vaccinia y variola: a study of their life history Churchill, Londres
- ↑ Stanley WM, Loring HS (1936). "The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants". Science, 83, p.85 PMID 17756690.
- ↑ Stanley WM, Lauffer MA (1939). "Disintegration of tobacco mosaic virus in urea solutions". Science 88, pp. 345–347 PMID 17788438
- ↑ Creager AN, Morgan GJ (Junio de 2008). "After the double helix: Rosalind Franklin's research on Tobacco mosaic virus". Isis 99 (2): 239–72. doi:10.1086/588626. PMID 18702397
- ↑ Dimmock p. 12
- ↑ Norrby E (2008). "Nobel Prizes and the emerging virus concept". Arch. Virol. 153 (6): 1109–23. doi:10.1007/s00705-008-0088-8. PMID 18446425
- ↑ Collier p. 745
- ↑ a b Temin HM, Baltimore D (1972). "RNA-directed DNA synthesis and RNA tumor viruses". Adv. Virus Res. 17: 129–86. doi:10.1016/S0065-3527(08)60749-6. PMID 4348509
- ↑ Barré-Sinoussi, F., Chermann, J. C., Rey, F., Nugeyre, M. T., Chamaret, S., Gruest, J., Dauguet, C., Axler-Blin, C., Vezinet-Brun, F., Rouzioux, C., Rozenbaum, W., and Montagnier, L. (1983). "Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS)". Science 220 (4599): 868–871. doi:10.1126/science.6189183. PMID 6189183.
- ↑ Institut de Génétique et Microbiologie (Abril 2006). «The origin of viruses and their possible roles in major evolutionary transitions.». Virus Res. 5 (117): pp. 5-16. PMID 16476498. http://www.ncbi.nlm.nih.gov/pubmed/16476498.
- ↑ Koonin EV, Senkevich TG, Dolja VV. (Septiembre 2006). «The ancient Virus World and evolution of cells.». PUBMED I: pp. 29. PMID 16984643. http://www.ncbi.nlm.nih.gov/pubmed/16984643.
- ↑ Iyer LM, Balaji S, Koonin EV, Aravind L (Abril 2006). «Evolutionary genomics of nucleo-cytoplasmic large DNA viruses». Virus Res. 117 (1): pp. 156–84. doi:. PMID 16494962. http://linkinghub.elsevier.com/retrieve/pii/S0168-1702(06)00028-1.
- ↑ Liu Y, Nickle DC, Shriner D, et al. (2004). "Molecular clock-like evolution of human immunodeficiency virus type 1". Virology. 10;329(1):101–8, PMID 15476878
- ↑ Shores p. 16
- ↑ Collier pp. 18-19
- ↑ Shores pp. 14-16
- ↑ Collier pp. 11-21
- ↑ a b Dimmock p. 16
- ↑ Collier, Balows y Sussman p. 11
- ↑ Collier pp. 11–12
- ↑ Shors p. 574
- ↑ McClintock, B. (Junio 1950). «The origin and behavior of mutable loci in maize». Proc Natl Acad Sci U S A. 36 (6): pp. 344–55. doi:. PMID 15430309.
- ↑ Dimmock p. 55
- ↑ Shors 551–3
- ↑ Tsagris EM, de Alba AE, Gozmanova M, Kalantidis K (Septiembre 2008). «Viroids». Cell. Microbiol. 10 (11): pp. 2168. doi:. PMID 18764915. http://www.blackwell-synergy.com/doi/abs/10.1111/j.1462-5822.2008.01231.x.
- ↑ Shors p. 492–3
- ↑ La Scola B, Desnues C, Pagnier I, Robert C, Barrassi L, Fournous G, Merchat M, Suzan-Monti M, Forterre P, Koonin E, Raoult D (Septiembre 2008). «The virophage as a unique parasite of the giant mimivirus». Nature 455 (7209): pp. 100–4. doi:. PMID 18690211.
- ↑ Collier p. 777
- ↑ Dimmock págs. 55–7
- ↑ Liberski PP (2008). «Prion diseases: a riddle wrapped in a mystery inside an enigma». Folia Neuropathol 46 (2): pp. 93–116. PMID 18587704. http://www.folianeuro.termedia.pl/showarticle.php?id=10625.
- ↑ Dimmock pp. 57–58
- ↑ Lupi O, Dadalti P, Cruz E, Goodheart C (2007). «Did the first virus self-assemble from self-replicating prion proteins and RNA?». Med. Hypotheses 69 (4): pp. 724–30. doi:. PMID 17512677. http://linkinghub.elsevier.com/retrieve/pii/S0306-9877(07)00261-7.
- ↑ Dimmock pp. 15–16
- ↑ Rybicki EP (1990) "The classification of organisms at the edge of life, or problems with virus systematics." S Aft J Sci 86:182–186
- ↑ Holmes EC (Octubre 2007). «Viral evolution in the genomic age». PLoS Biol. 5 (10): pp. e278. doi:. PMID 17914905. PMC 1994994. http://biology.plosjournals.org/perlserv/?request=get-document&doi=10.1371/journal.pbio.0050278.
- ↑ Koonin EV, Senkevich TG, Dolja VV (2006). «The ancient Virus World and evolution of cells». Biol. Direct 1: p. 29 pp. 29. doi:. PMID 16984643. PMC 1594570. http://www.biology-direct.com/content/1//29.
- ↑ Keith Roberts, Martin Raff, Bruce Alberts, Peter Walter, Julian Lewis and Alexander Johnson, Molecular Biology of the Cell 4th Edition, Routledge, March, 2002, hardcover, 1616 pages, 7.6 pounds, ISBN 0-8153-3218-1
- ↑ ARANDA ANZALDO, Armando. La estructura de los virus. 1ro.. pp. 6-7. http://bibliotecadigital.ilce.edu.mx/sites/ciencia/volumen2/ciencia3/071/htm/sec_6.htm. Consultado el 26 de junio de 2010.
- ↑ a b c d e Calendar, R., The Bacteriophages, Dos volúmenes, (comp), Plenum Press, 1988.
- ↑ a b Collier pp. 33–55
- ↑ Collier pp. 33–37
- ↑ Kiselev NA, Sherman MB, Tsuprun VL (1990). «Negative staining of proteins». Electron Microsc. Rev. 3 (1): pp. 43–72. doi:. PMID 1715774.
- ↑ Collier p. 40
- ↑ Caspar DL, Klug A (1962). «Physical principles in the construction of regular viruses». Cold Spring Harb. Symp. Quant. Biol. 27: pp. 1–24. PMID 14019094.
- ↑ Crick FH, Watson JD (1956). «Structure of small viruses». Nature 177 (4506): pp. 473–5. doi:. PMID 13309339.
- ↑ Falvo, M.R.; S. Washburn, R. Superfine, M. Finch, F.P. Brooks Jr, V. Chi, R.M. Taylor 2nd (Marzo de 1997). «Manipulation of individual viruses: friction and mechanical properties». Biophysical Journal 72 (3): pp. 1396–1403. doi:. PMID 9138585. PMC 1184522. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B94RW-4V9K7FD-1D&_user=108429&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000059713&_version=1&_urlVersion=0&_userid=108429&md5=d5aafeb39c5a05782d0935dc7ced27ad.
- ↑ Kuznetsov, Yu. G.; A. J. Malkin, R. W. Lucas, M. Plomp, A. McPherson (1 de septiembre de 2001). «Imaging of viruses by atomic force microscopy». J Gen Virol 82 (9): pp. 2025–2034. http://vir.sgmjournals.org.
- ↑ Collier p. 37
- ↑ Collier pp. 40, 42
- ↑ Collier pp. 42–43
- ↑ Rossmann MG, Mesyanzhinov VV, Arisaka F, Leiman PG (Abril 2004). «The bacteriophage T4 DNA injection machine». Curr. Opin. Struct. Biol. 14 (2): pp. 171–80. doi:. PMID 15093831.
- ↑ Long GW, Nobel J, Murphy FA, Herrmann KL, Lourie B (Septiembre 1970). «Experience with electron microscopy in the differential diagnosis of smallpox». Appl Microbiol 20 (3): pp. 497–504. PMID 4322005.
- ↑ Suzan-Monti M, La Scola B, Raoult D (2006). «Genomic and evolutionary aspects of Mimivirus». Virus Research 117 (1): pp. 145–155. doi:. PMID 16181700.
- ↑ a b Prangishvili D, Forterre P, Garrett RA (2006). «Viruses of the Archaea: a unifying view». Nat. Rev. Microbiol. 4 (11): pp. 837–48. doi:. PMID 17041631.
- ↑ a b c d Collier pp. 96–99
- ↑ Pressing J, Reanney DC (1984). "Divided genomes and intrinsic noise". J Mol Evol. 20(2):135–46.
- ↑ Pan XP, Li LJ, Du WB, Li MW, Cao HC, Sheng JF (Noviembre 2007). «Differences of YMDD mutational patterns, precore/core promoter mutations, serum HBV DNA levels in lamivudine-resistant hepatitis B genotypes B and C». J. Viral Hepat. 14 (11): pp. 767–74. doi:. PMID 17927612. http://www.blackwell-synergy.com/openurl?genre=article&sid=nlm:pubmed&issn=1352-0504&fecha=2007&volumen=14&número=11&spage=767.
- ↑ Hampson AW, Mackenzie JS (Noviembre 2006). «The influenza viruses». Med. J. Aust. 185 (10 Suppl): pp. S39–43. PMID 17115950. http://www.mja.com.au/public/issues/185_10_201106/ham10884_fm.html.
- ↑ Metzner KJ (Diciembre 2006). «Detection and significance of minority quasispecies of drug-resistant HIV-1». J HIV Ther 11 (4): pp. 74–81. PMID 17578210.
- ↑ Goudsmit, Jaap. Viral Sex. Oxford Univ Press, 1998.ISBN 978-0-19-512496-5 ISBN 0-19-512496-0
- ↑ Worobey M, Holmes EC (1999). «Evolutionary aspects of recombination in RNA viruses». J. Gen. Virol. 80 ( Pt 10): pp. 2535–43. PMID 10573145.
- ↑ Lukashev AN (2005). «Role of recombination in evolution of enteroviruses». Rev. Med. Virol. 15 (3): pp. 157–67. doi:. PMID 15578739.
- ↑ Umene K (1999). «Mechanism and application of genetic recombination in herpesviruses». Rev. Med. Virol. 9 (3): pp. 171–82. doi:. PMID 10479778.
- ↑ Dimmock p. 70
- ↑ Boevink P, Oparka KJ (Agosto 2005). «Virus-host interactions during movement processes». Plant Physiol. 138 (4): pp. 1815–21. doi:. PMID 16172094. PMC 1183373. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=16172094.
- ↑ Dimmock p. 71
- ↑ Barman S, Ali A, Hui EK, Adhikary L, Nayak DP (2001). «Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses». Virus Res. 77 (1): pp. 61–9. doi:. PMID 11451488.
- ↑ Shors p. 54; Collier p. 78
- ↑ a b N.J. Dimmock, A.J. Easton y K. Leppard, Introduction to Modern Virology, Ed. Wiley, 2006, ISBN 978-1-4051-3645-7.
- ↑ a b Robertson MP, Igel H, Baertsch R, Haussler D, Ares M Jr, Scott WG (2005). «The structure of a rigorously conserved RNA element within the SARS virus genome». PLoS Biol 3 (1): pp. e5. PMID 15630477 doi 10.1371/journal.pbio.0030005.
- ↑ John Carter y Venetia Saunders, Virology: Principles and Applications, Ed. Wiley, 2007, ISBN 978-0-470-02387-7.
- ↑ a b c d e f g A. Kornberg y T.A Baker, DNA Replication, Ed. University Science Books, 2005, ISBN 978-1-891389-44-3.
- ↑ Collier p. 79
- ↑ Collier pp. 115–146
- ↑ Collier p. 115
- ↑ Roulston A, Marcellus RC, Branton PE (1999). «Viruses and apoptosis». Annu. Rev. Microbiol. 53: pp. 577–628. doi:. PMID 10547702. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.micro.53.1.577?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%3dncbi.nlm.nih.gov.
- ↑ Alwine JC (2008). «Modulation of host cell stress responses by human cytomegalovirus». Curr. Top. Microbiol. Immunol. 325: pp. 263–79. doi:. PMID 18637511.
- ↑ Sinclair J (Marzo 2008). «Human cytomegalovirus: Latency and reactivation in the myeloid lineage». J. Clin. Virol. 41 (3): pp. 180–5. doi:. PMID 18164651. http://linkinghub.elsevier.com/retrieve/pii/S1386-6532(07)00434-9.
- ↑ Jordan MC, Jordan GW, Stevens JG, Miller G (Junio 1984). «Latent herpesviruses of humans». Ann. Intern. Med. 100 (6): pp. 866–80. PMID 6326635.
- ↑ Sissons JG, Bain M, Wills MR (Febrero 2002). «Latency and reactivation of human cytomegalovirus». J. Infect. 44 (2): pp. 73–7. doi:. PMID 12076064. http://linkinghub.elsevier.com/retrieve/pii/S016344530190948X.
- ↑ Barozzi P, Potenza L, Riva G, Vallerini D, Quadrelli C, Bosco R, Forghieri F, Torelli G, Luppi M (Diciembre 2007). «B cells and herpesviruses: a model of lymphoproliferation». Autoimmun Rev 7 (2): pp. 132–6. doi:. PMID 18035323. http://linkinghub.elsevier.com/retrieve/pii/S1568-9972(07)00061-4.
- ↑ Subramanya D, Grivas PD (Noviembre 2008). «HPV and cervical cancer: updates on an established relationship». Postgrad Med 120 (4): pp. 7–13. doi:. PMID 19020360.
- ↑ Lwoff A, Horne RW, Tournier P (1962). «A virus system» (en French). C. R. Hebd. Seances Acad. Sci. 254: pp. 4225–7. PMID 14467544.
- ↑ Lwoff A, Horne R, Tournier P (1962). «A system of viruses». Cold Spring Harb. Symp. Quant. Biol. 27: pp. 51–5. PMID 13931895.
- ↑ Fields p. 27
- ↑ Delwart EL (2007). «Viral metagenomics». Rev. Med. Virol. 17 (2): pp. 115–31. doi:. PMID 17295196.
- ↑ Virus Taxonomy 2008. International Committee on Taxonomy of Viruses. Consultado el 15 de septiembre de 2008.
- ↑ ICTV Master Species List 2008
- Archivo con reportes. Producido por el Comité Internacional sobre Taxonomía de Virus. Consultado el 15 de septiembre de 2008
- ↑ Baltimore D (1974). «The strategy of RNA viruses». Harvey Lect. 70 Series: pp. 57–74. PMID 4377923.
- ↑ van Regenmortel MH, Mahy BW (2004). «Emerging issues in virus taxonomy». Emerging Infect. Dis. 10 (1): pp. 8–13. PMID 15078590.
- ↑ Mayo MA (1999). «Developments in plant virus taxonomy since the publication of the 6th ICTV Report. International Committee on Taxonomy of Viruses». Arch. Virol. 144 (8): pp. 1659–66. doi:. PMID 10486120.
- ↑ de Villiers EM, Fauquet C, Broker TR, Bernard HU, zur Hausen H (2004). «Classification of papillomaviruses». Virology 324 (1): pp. 17–27. doi:. PMID 15183049.
- ↑ Mainly Chapter 33 (Disease summaries), págs. 367–392 en:Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. (2007). Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series). Hagerstwon, MD: Lippincott Williams & Wilkins. pp. págs. 367–392. ISBN 0-7817-8215-5.|alt=A photograph of the upper body of a man labelled with the names of viruses that infect the different parts
- ↑ For common cold: National Institute of Allergy and Infectious Diseases (NIAID) > Common Cold. Última modificación 10 de diciembre de 2007. Consultado el 4 de abril 2009
- ↑ Thiel V (editor). (2007). Coronaviruses: Molecular and Cellular Biology (1st edición). Caister Academic Press. ISBN [[Especial:FuentesDeLibros/978-1-904455-16-5]|978-1-904455-16-5]]]. http://www.horizonpress.com/cor.
- ↑ Akhtar, Jihan; Shukla, Deepak. «Viral entry mechanisms: cellular and viral mediators of herpes simplex virus entry». FEBS Journal 2009 (276). http://www3.interscience.wiley.com/cgi-bin/fulltext/122668866/PDFSTART.
- ↑ Kawaoka Y (editor). (2006). Influenza Virology: Current Topics. Caister Academic Press. ISBN 978-1-904455-06-6. http://www.horizonpress.com/flu.
- ↑ Smith DK, Grohskopf LA, Black RJ, et al. (2005). «Antiretroviral Postexposure Prophylaxis After Sexual, Injection-Drug Use, or Other Nonoccupational Exposure to HIV in the United States». MMWR 54 (RR02): pp. 1–20. http://www.cdc.gov/mmwr/preview/mmwrhtml/rr5402a1.htm#tab1.
- ↑ Thacker E, Janke B (February 2008). «Swine influenza virus: zoonotic potential and vaccination strategies for the control of avian and swine influenzas». The Journal of Infectious Diseases 197 Suppl 1: pp. S19–24. doi:. PMID 18269323.
- ↑ Klenk, Hans-Dieter; Feldmann, Heinz (2004). Ebola and Marburg Viruses, Molecular and Cellular Biology, Wymondham, Norfolk: Horizon Bioscience. ISBN 0-9545232-3-7.
- ↑ Komaroff AL (Diciembre 2006). «Is human herpesvirus-6 a trigger for chronic fatigue syndrome?». J. Clin. Virol. 37 Suppl 1: pp. S39–46. doi:. PMID 17276367. http://www.hhv-6foundation.org/hhv6cfs_komaroff.html.
- ↑ Chen C, Chiu Y, Wei F, Koong F, Liu H, Shaw C, Hwu H, Hsiao K (1999). «High seroprevalence of Borna virus infection in schizophrenic patients, family members and mental health workers in Taiwan». Mol Psychiatry 4 (1): pp. 33–8. doi:. PMID 10089006.
- ↑ Margolis TP, Elfman FL, Leib D, et al. (Octubre 2007). «Spontaneous reactivation of herpes simplex virus type 1 in latently infected murine sensory ganglia». J. Virol. 81 (20): pp. 11069–74. doi:. PMID 17686862. PMC 2045564. http://jvi.asm.org/cgi/pmidlookup?view=long&pmid=17686862.
- ↑ Whitley RJ, Roizman B (Mayo 2001). «Herpes simplex virus infections». Lancet 357 (9267): pp. 1513–8. doi:. PMID 11377626.
- ↑ Barton ES, White DW, Cathelyn JS, et al. (Mayo 2007). «Herpesvirus latency confers symbiotic protection from bacterial infection». Nature 447 (7142): pp. 326–9. doi:. PMID 17507983.
- ↑ Bertoletti A, Gehring A (2007). «Immune response and tolerance during chronic hepatitis B virus infection». Hepatol. Res. 37 Suppl 3: pp. S331–8. doi:. PMID 17931183.
- ↑ Rodrigues C, Deshmukh M, Jacob T, Nukala R, Menon S, Mehta A (2001). «Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients». Indian journal of medical microbiology 19 (3): pp. 141–4. PMID 17664817.
- ↑ Nguyen VT, McLaws ML, Dore GJ (2007). «Highly endemic hepatitis B infection in rural Vietnam». Journal of Gastroenterology and Hepatology 22 (12): p. 2093 pp. 2093–100. doi:. PMID 17645465.
- ↑ Witzany G (2006) "Natural Genome-Editing Competences of Viruses'" Acta Biotheor 54: 235–253.
- ↑ Fowler MG, Lampe MA, Jamieson DJ, Kourtis AP, Rogers MF (2007). «Reducing the risk of mother-to-child human immunodeficiency virus transmission: past successes, current progress and challenges, and future directions». Am. J. Obstet. Gynecol. 197 (3 Suppl): pp. S3–9. doi:. PMID 17825648.
- ↑ Sauerbrei A, Wutzler P (2000). «The congenital varicella syndrome». Journal of perinatology : official journal of the California Perinatal Association 20 (8 Pt 1): pp. 548–54. PMID 11190597.
- ↑ Garnett GP (Febrero 2005). «Role of herd immunity in determining the effect of vaccines against sexually transmitted disease». J. Infect. Dis. 191 Suppl 1: pp. S97–106. doi:. PMID 15627236. http://www.journals.uchicago.edu/cgi-bin/resolve?JID31548.
- ↑ Platonov AE (2006). «(The influence of weather conditions on the epidemiology of vector-borne diseases by the example of West Nile fever in Russia)» (en ruso). Vestn. Akad. Med. Nauk SSSR (2): pp. 25–9. PMID 16544901.
- ↑ Shors p. 198
- ↑ Shors pp. 199, 209
- ↑ Shors p. 19
- ↑ Shors p. 126
- ↑ Shors pp. 193–194
- ↑ Shors pp. 193–94
- ↑ Shors p. 194
- ↑ Shors pp. 192–193
- ↑ Ranlet P (2000). «The British, the Indians, and smallpox: what actually happened at Fort Pitt in 1763?». Pa Hist 67 (3): pp. 427–41. PMID 17216901.
- ↑ Van Rijn K (2006). «"Lo! The poor Indian!" colonial responses to the 1862–63 smallpox epidemic in British Columbia and Vancouver Island». Can Bull Med Hist 23 (2): pp. 541–60. PMID 17214129.
- ↑ Patterson KB, Runge T (Abril 2002). «Smallpox and the Native American». Am. J. Med. Sci. 323 (4): pp. 216–22. doi:. PMID 12003378.
- ↑ Sessa R, Palagiano C, Scifoni MG, di Pietro M, Del Piano M (Marzo 1999). «The major epidemic infections: a gift from the Old World to the New?». Panminerva Med 41 (1): pp. 78–84. PMID 10230264.
- ↑ Bianchine PJ, Russo TA (1992). «The role of epidemic infectious diseases in the discovery of America». Allergy Proc 13 (5): pp. 225–32. doi:. PMID 1483570. http://openurl.ingenta.com/content/nlm?genre=article&issn=1088-5412&volumen=13&número=5&spage=225&aulast=Bianchine.
- ↑ Hauptman LM (Noviembre 1979). «Smallpox and American Indian; Depopulation in Colonial New York». N Y State J Med 79 (12): pp. 1945–9. PMID 390434.
- ↑ Fortuine R (1988). «Smallpox decimates the Tlingit (1787)». Alaska Med 30 (3): p. 109 pp. 109. PMID 3041871.
- ↑ Collier pp. 409–415
- ↑ Patterson KD, Pyle GF (Spring 1991). «The geography and mortality of the 1918 influenza pandemic». Bull Hist Med. 65 (1): pp. 4–21. PMID 2021692.
- ↑ Johnson NP, Mueller J (2002). «Updating the accounts: global mortality of the 1918–1920 "Spanish" influenza pandemic». Bull Hist Med 76 (1): pp. 105–15. doi:. PMID 11875246. http://muse.jhu.edu/cgi-bin/resolve_openurl.cgi?issn=0007-5140&volumen=76&número=1&spage=105&aulast=Johnson.
- ↑ Gao F, Bailes E, Robertson DL, et al. (1999). «Origin of HIV-1 in the Chimpanzee Pan troglodytes troglodytes». Nature 397 (6718): pp. 436–441. doi:. PMID doi [http://dx.doi.org/10.1038/17130 10.1038/17130 9989410 doi 10.1038/17130].
- ↑ Shors p. 447
- ↑ Mawar N, Saha S, Pandit A, Mahajan U (Diciembre 2005). «The third phase of HIV pandemic: social consequences of HIV/AIDS stigma & discrimination & future needs» (PDF). Indian J. Med. Res. 122 (6): pp. 471–84. PMID 16517997. http://www.icmr.nic.in/ijmr/2005/december/1201.pdf.
- ↑ «Status of the global HIV epidemic» (PDF). UNAIDS (2008). Consultado el 15 de septiembre de 2008.
- ↑ Towner JS, Khristova ML, Sealy TK, et al. (Julio 2006). «Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola». J. Virol. 80 (13): pp. 6497–516. doi:. PMID 16775337. PMC 1488971. http://jvi.asm.org/cgi/pmidlookup?view=long&pmid=16775337.
- ↑ «Brotes de gripe en México y en los Estados Unidos». OMS/WHO (abril de 2009). Consultado el 26 de abril de 2009. «La OMS emite su opinión en cuanto al brote de gripe porcina del 2009.».
- ↑ «Press Release: A/H1N1 influenza like human illness in Mexico and the USA: OIE statement». World Organisation for Animal Health (29 de abril de 2009). Consultado el 29 de abril de 2009.
- ↑ «OMS: Gripe porcina tiene potencial pandémico». RFI (25 de abril de 2009). Consultado el 26 de abril de 2009.
- ↑ «Reporte diario del ECDC sobre la pandemia de influenza A (H1N1) de 2009» (en inglés). Centro Europeo para la Prevención y Control de Enfermedades (18-01-2010 09:00 UTC +2).
- ↑ Einstein MH, Schiller JT, Viscidi RP, Strickler HD, Coursaget P, Tan T, Halsey N, Jenkins D (Junio 2009). «Clinician's guide to human papillomavirus immunology: knowns and unknowns». The Lancet Infectious Diseases 9 (6): pp. 347–56. doi:. PMID 19467474.
- ↑ Shuda M, Feng H, Kwun HJ, Rosen ST, Gjoerup O, Moore PS, Chang Y (Octubre 2008). «T antigen mutations are a human tumor-specific signature for Merkel cell polyomavirus». Proceedings of the National Academy of Sciences of the United States of America 105 (42): pp. 16272–7. doi:. PMID 18812503.
- ↑ Pulitzer MP, Amin BD, Busam KJ (Mayo 2009). «Merkel cell carcinoma: review». Advances in Anatomic Pathology 16 (3): pp. 135–44. doi:. PMID 19395876.
- ↑ Koike K (2007). «Hepatitis C virus contributes to hepatocarcinogenesis by modulating metabolic and intracellular signalling pathways». J. Gastroenterol. Hepatol. 22 Suppl 1: pp. S108–11. doi:. PMID 17567457.
- ↑ Hu J, Ludgate L (2007). «HIV-HBV and HIV-HCV coinfection and liver cancer development». Cancer Treat. Res. 133: pp. 241–52. doi:. PMID 17672044.
- ↑ Bellon M, Nicot C (2007). «Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia». Cancer genomics & proteomics 4 (1): pp. 21–5. PMID 17726237.
- ↑ Schiffman M, Castle PE, Jeronimo J, Rodriguez AC, Wacholder S (2007). «Human papillomavirus and cervical cancer». Lancet 370 (9590): pp. 890–907. doi:. PMID 17826171.
- ↑ Klein E, Kis LL, Klein G (2007). «Epstein-Barr virus infection in humans: from harmless to life endangering virus-lymphocyte interactions». Oncogene 26 (9): pp. 1297–305. doi:. PMID 17322915.
- ↑ zur Hausen H (Julio 2008). «Novel human polyomaviruses—re-emergence of a well known virus family as possible human carcinogens». International Journal of Cancer. Journal International Du Cancer 123 (2): pp. 247–50. doi:. PMID 18449881.
- ↑ Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?call=bv.View..ShowTOC&rid=mboc4.TOC&depth=2.
- ↑ Ding SW, Voinnet O. Antiviral immunity directed by small RNAs. Cell. 2007 Aug 10;130(3):413-26.
- ↑ Patton JT, Vasquez-Del Carpio R, Spencer E (2004). «Replication and transcription of the rotavirus genome». Curr. Pharm. Des. 10 (30): pp. 3769–77. doi:. PMID 15579070.
- ↑ Jayaram H, Estes MK, Prasad BV (2004). «Emerging themes in rotavirus cell entry, genome organization, transcription and replication». Virus Res. 101 (1): pp. 67–81. doi:. PMID 15010218.
- ↑ Greer S, Alexander GJ. Viral serology and detection. Baillieres Clin Gastroenterol. 1995 Dec;9(4):689-721
- ↑ Laurence JC. Hepatitis A and B immunizations of individuals infected with humanimmunodeficiency virus.Am J Med. 2005 Oct;118 Suppl 10A:75S-83S.
- ↑ Cascalho M, Platt JL. Novel functions of B cells.Crit Rev Immunol. 2007;27(2):141-51.
- ↑ Khatri M, Sharma JM. Modulation of macrophages by infectious bursal disease virus.Cytogenet Genome Res. 2007;117(1-4):388-93
- ↑ Le Page C, Genin P, Baines MG, Hiscott J. Interferon activation and innate immunity.Rev Immunogenet. 2000;2(3):374-86.
- ↑ Hilleman MR. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections. Proc Natl Acad Sci U S A. 2004 Oct 5;101 Suppl 2:14560-6. Epub 2004 Aug 5.
- ↑ Asaria P, MacMahon E (2006). «Measles in the United Kingdom: can we eradicate it by 2010?». BMJ 333 (7574): pp. 890–5. doi:. PMID 17068034.
- ↑ Lane JM (2006). «Mass vaccination and surveillance/containment in the eradication of smallpox». Curr. Top. Microbiol. Immunol. 304: pp. 17–29. PMID 16989262.
- ↑ Arvin AM, Greenberg HB (2006). «New viral vaccines». Virology 344 (1): pp. 240–9. doi:. PMID 16364754.
- ↑ Pastoret PP, Schudel AA, Lombard M (2007). «Conclusions--future trends in veterinary vaccinology». Rev. - Off. Int. Epizoot. 26 (2): pp. 489–94, 495–501, 503–9. PMID 17892169.
- ↑ Mayer, Gene (2006). «Immunology - Chapter One: Innate (non-specific) Immunity». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Consultado el 1 de enero de 2007.
- ↑ Litman G, Cannon J, Dishaw L (2005). «Reconstructing immune phylogeny: new perspectives.». Nat Rev Immunol 5 (11): pp. 866-79. PMID 16261174.
- ↑ Palese P (2006). «Making better influenza virus vaccines?». Emerging Infect. Dis. 12 (1): pp. 61–5. PMID 16494719.
- ↑ Thomssen R (1975). «Live attenuated versus killed virus vaccines». Monographs in allergy 9: pp. 155–76. PMID 1090805.
- ↑ McLean AA (1986). «Development of vaccines against hepatitis A and hepatitis B». Rev. Infect. Dis. 8 (4): pp. 591–8. PMID 3018891.
- ↑ Casswall TH, Fischler B (2005). «Vaccination of the immunocompromised child». Expert review of vaccines 4 (5): pp. 725–38. doi:. PMID 16221073.
- ↑ «A history of de Discovery and Clinical Application of Antiviral». British Medical Bulletin (41): pp. 309-314. 1985. http://bmb.oxfordjournals.org/cgi/content/abstract/41/4/309.
- ↑ Irwin, MR., Olmstead, R., Oxman, MN. (2007). Augmenting Immune Responses to Varicella Zoster Virus in Older Adults: A Randomized, Controlled Trial of Tai Chi. J Am Geriatr Soc 55 (4): 511-517. ISSN 1532-5415. Último acceso el 6 de septiembre de 2007. En inglés.
- ↑ Magden J, Kääriäinen L, Ahola T (2005). «Inhibitors of virus replication: recent developments and prospects». Appl. Microbiol. Biotechnol. 66 (6): pp. 612–21. doi:. PMID 15592828.
- ↑ Mindel A, Sutherland S (1983). «Genital herpes - the disease and its treatment including intravenous acyclovir». J. Antimicrob. Chemother. 12 Suppl B: pp. 51–9. PMID 6355051.
- ↑ Witthoft T, Moller B, Wiedmann KH, Mauss S, Link R, Lohmeyer J, Lafrenz M,Gelbmann CM, Huppe D, Niederau C, Alshuth U. Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice: The German Open Safety Trial. J Viral Hepat. 2007 Nov;14(11):788-96.
- ↑ Rudin D, Shah SM, Kiss A, Wetz RV, Sottile VM. Interferon and lamivudine vs. interferon for hepatitis B e antigen-positive hepatitis B treatment: meta-analysis of randomized controlled trials.Liver Int. 2007 Nov;27(9):1185-93.
- ↑ Dimmock p. 3
- ↑ Bernard La Scola, Christelle Desnues, Isabelle Pagnier, Catherine Robert, Lina Barrassi, Ghislain Fournous, Michèle Merchat, Marie Suzan-Monti, Patrick Forterre, Eugene Koonin and Didier Raoult (2008). «The virophage as a unique parasite of the giant mimivirus». Nature 454 (7205): pp. 100. doi:.
- ↑ Goris N, Vandenbussche F, De Clercq K (Abril 2008). «Potential of antiviral therapy and prophylaxis for controlling RNA viral infections of livestock». Antiviral Res. 78 (1): pp. 170–8. doi:. PMID 18035428. http://linkinghub.elsevier.com/retrieve/pii/S0166-3542(07)00435-4.
- ↑ Carmichael L (2005). «An annotated historical account of canine parvovirus». J. Vet. Med. B Infect. Dis. Vet. Public Health 52 (7–8): pp. 303–11. doi:. PMID 16316389.
- ↑ Chen YP, Zhao Y, Hammond J, Hsu H, Evans JD, Feldlaufer MF (Octubre–Noviembre de 2004). «Multiple virus infections in the honey bee and genome divergence of honey bee viruses». Journal of Invertebrate Pathology 87 (2–3): pp. 84–93. doi:. PMID 15579317.
- ↑ Shors p. 584
- ↑ Shors pp. 562–587
- ↑ Dinesh-Kumar SP, Wai-Hong Tham, Baker BJ (2000). "Structure—function analysis of the tobacco mosaic virus resistance gene N". PNAS 97, 14789-94 PMID 11121079
- ↑ Soosaar JL, Burch-Smith TM, Dinesh-Kumar SP (2005). "Mechanisms of plant resistance to viruses". Nat. Rev. Microbiol. 3, pp. 789–98 PMID 16132037
- ↑ Wommack KE, Colwell RR (Marzo 2000). «Virioplankton: viruses in aquatic ecosystems». Microbiol. Mol. Biol. Rev. 64 (1): pp. 69–114. doi:. PMID 10704475. PMC 98987. http://mmbr.asm.org/cgi/pmidlookup?view=long&pmid=10704475.
- ↑ Bergh O, Børsheim KY, Bratbak G, Heldal M (Agosto 1989). «High abundance of viruses found in aquatic environments». Nature 340 (6233): pp. 467–8. doi:. PMID 2755508.
- ↑ Shors pp. 595–97
- ↑ Bickle TA, Krüger DH (1 de junio de 1993). «Biology of DNA restriction». Microbiol. Rev. 57 (2): pp. 434–50. PMID 8336674. PMC 372918. http://mmbr.asm.org/cgi/pmidlookup?view=long&pmid=8336674.
- ↑ Barrangou R, Fremaux C, Deveau H, et al. (Marzo 2007). «CRISPR provides acquired resistance against viruses in prokaryotes». Science (journal) 315 (5819): pp. 1709–12. doi:. PMID 17379808.
- ↑ Brouns SJ, Jore MM, Lundgren M, et al. (Agosto 2008). «Small CRISPR RNAs guide antiviral defense in prokaryotes». Science (journal) 321 (5891): pp. 960–4. doi:. PMID 18703739.
- ↑ Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E (2005). «Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements». J. Mol. Evol. 60 (2): pp. 174–82. doi:. PMID 15791728.
- ↑ Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV (2006). «A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action». Biol. Direct 1: p. 7 pp. 7. doi:. PMID 16545108. PMC 1462988. http://www.biology-direct.com/content/1/1/7.
- ↑ Collier p.8
- ↑ Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James.Viruses:Structure, Function, and Uses Consultado el 16 de septiembre de 2008
- ↑ Matsuzaki S, Rashel M, Uchiyama J, et al. (2005). «Bacteriophage therapy: a revitalized therapy against bacterial infectious diseases». J. Infect. Chemother. 11 (5): pp. 211–9. doi:. PMID 16258815.
- ↑ Fischlechner M, Donath E, M; Donath, E (2007). «Viruses as Building Blocks for Materials and Devices». Angewandte Chemie International Edition 46 (18): p. 3184 pp. 3184–93. doi:. PMID 17348058.
- ↑ Soto CM, Blum AS, Vora GJ, et al. (Abril 2006). «Fluorescent signal amplification of carbocyanine dyes using engineered viral nanoparticles». J. Am. Chem. Soc. 128 (15): pp. 5184–9. doi:. PMID 16608355.
- ↑ Blum AS, Soto CM, Wilson CD, et al. (2005). "An Engineered Virus as a Scaffold for Three-Dimensional Self-Assembly on the Nanoscale". Small, 7, 702.
- ↑ Shors p. 331
- ↑ a b c Artenstein AW, Grabenstein JD (Octubre 2008). «Smallpox vaccines for biodefense: need and feasibility». Expert Review of Vaccines 7 (8): pp. 1225–37. doi:. PMID 18844596. http://www.future-drugs.com/doi/abs/10.1586/14760584.7.8.1225?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%3dncbi.nlm.nih.gov.
- ↑ Aragón TJ, Ulrich S, Fernyak S, Rutherford GW (2003). «Risks of serious complications and death from smallpox vaccination: a systematic review of the United States experience, 1963–1968». BMC public health 3: p. 26 pp. 26. doi:. PMID 12911836.
- ↑ Weiss MM, Weiss PD, Mathisen G, Guze P (2004). «Rethinking smallpox». Clin. Infect. Dis. 39 (11): pp. 1668–73. doi:. PMID 15578369.
Bibliografía
- Collier, Leslie; Balows, Albert; Sussman, Max (1998) Topley and Wilson's Microbiology and Microbial Infections ninth edition, Volume 1, Virology, volume editors: Mahy, Brian and Collier, Leslie. Arnold. ISBN 0-340-66316-2.
- Dimmock, N.J; Easton, Andrew J; Leppard, Keith (2007) Introduction to Modern Virology sixth edition, Blackwell Publishing, ISBN 1-4051-3645-6.
- Knipe, David M; Howley, Peter M; Griffin, Diane E; Lamb, Robert A; Martin, Malcolm A; Roizman, Bernard; Straus Stephen E. (2007) Fields Virology, Lippincott Williams & Wilkins. ISBN 0-7817-6060-7.
- Shors, Teri (2008). Understanding Viruses. Jones and Bartlett Publishers. ISBN 0-7637-2932-9.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Virus. Commons
Wikimedia Commons alberga contenido multimedia sobre Virus. Commons Wikiespecies tiene un artículo sobre Virus. Wikispecies
Wikiespecies tiene un artículo sobre Virus. Wikispecies Wikcionario tiene definiciones para virus.Wikcionario
Wikcionario tiene definiciones para virus.Wikcionario

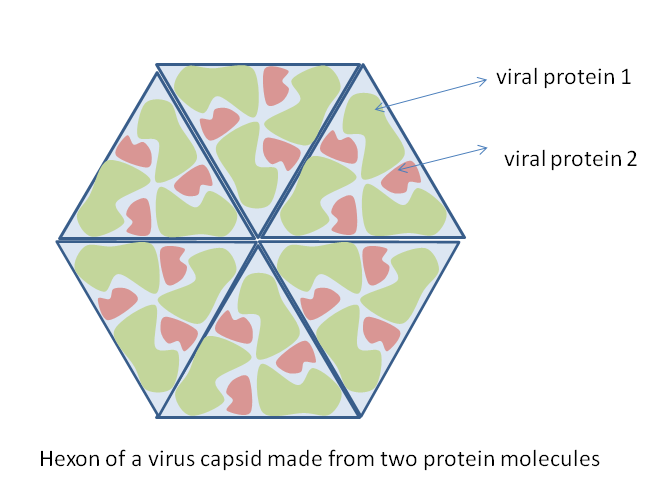

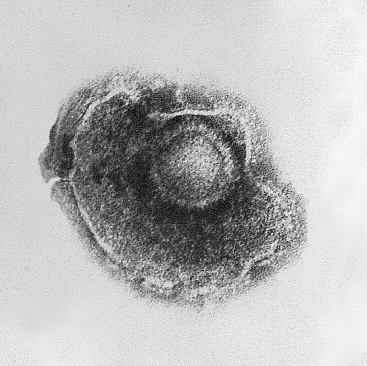
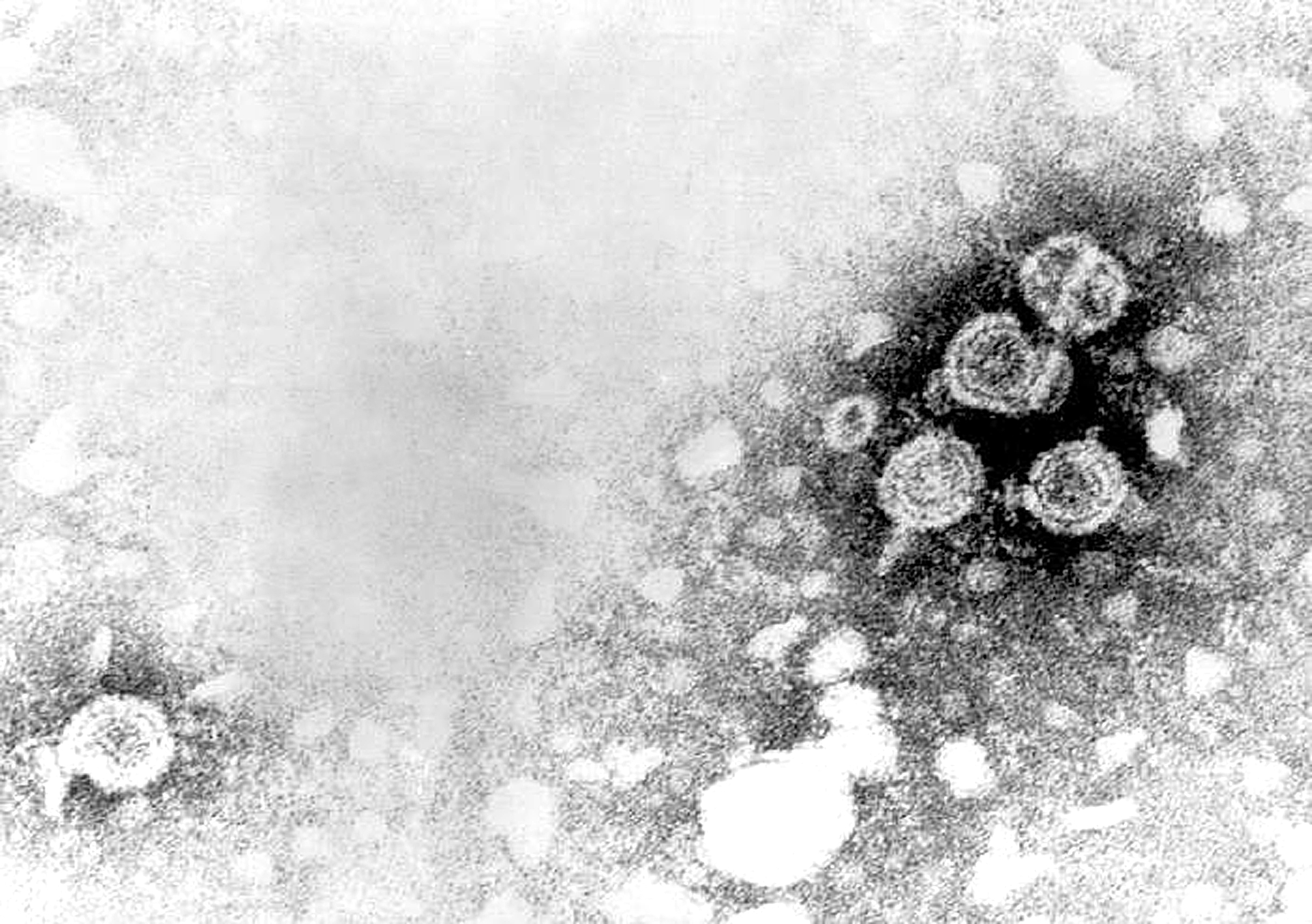
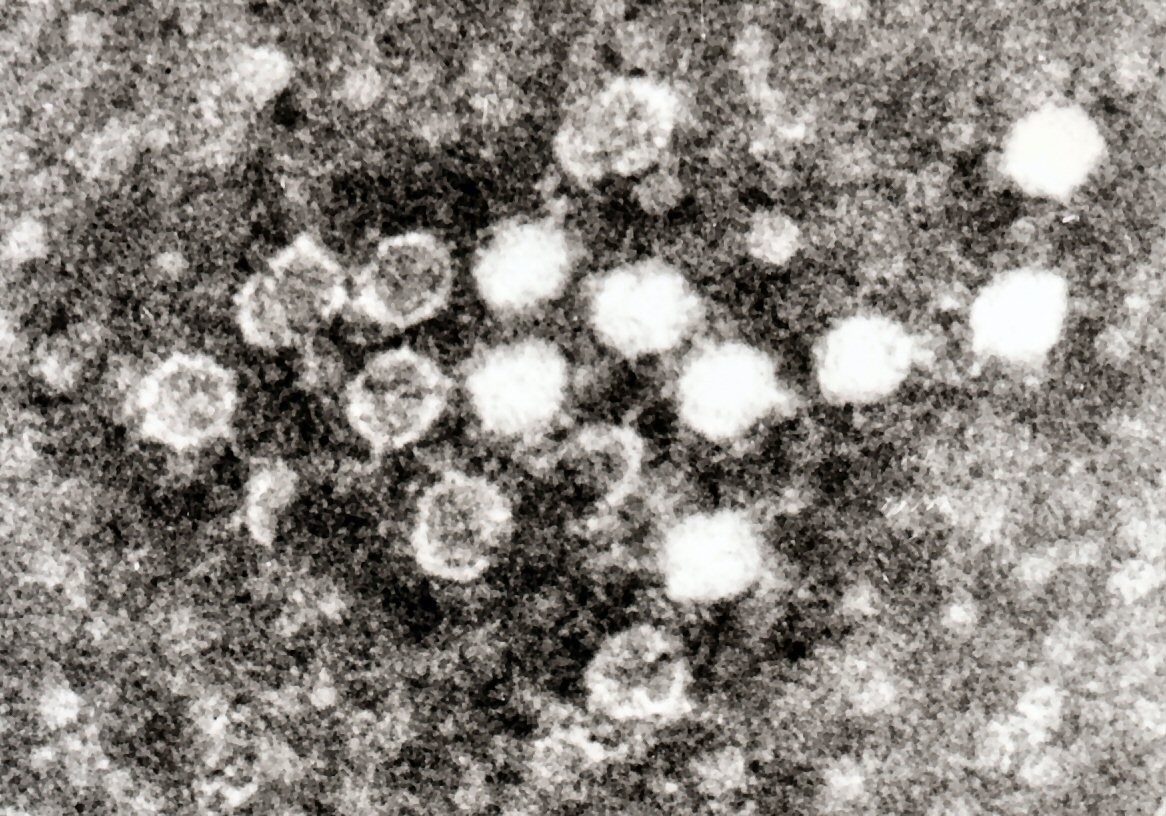
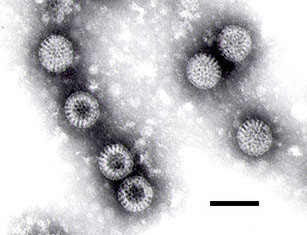
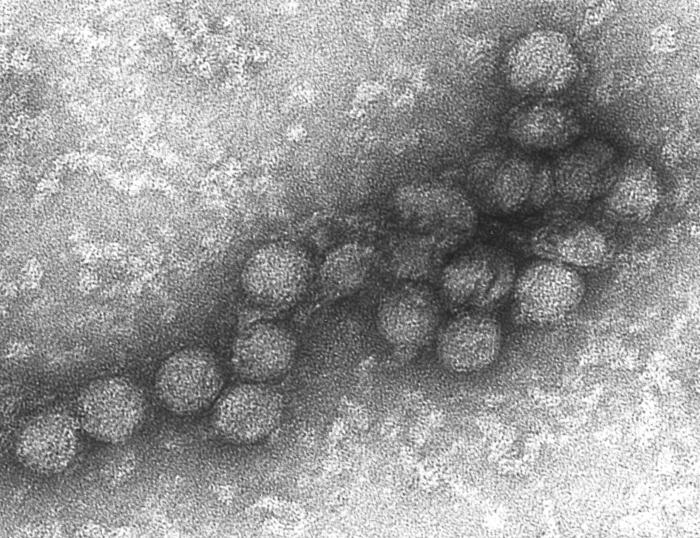
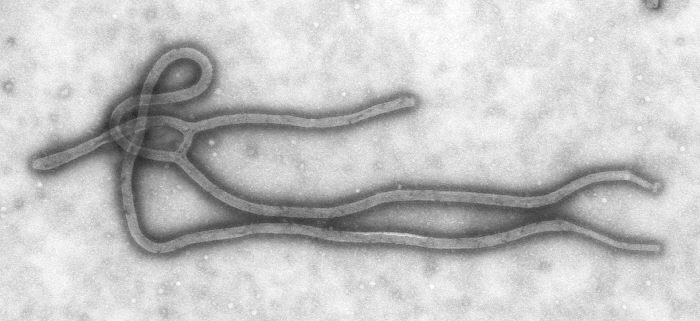

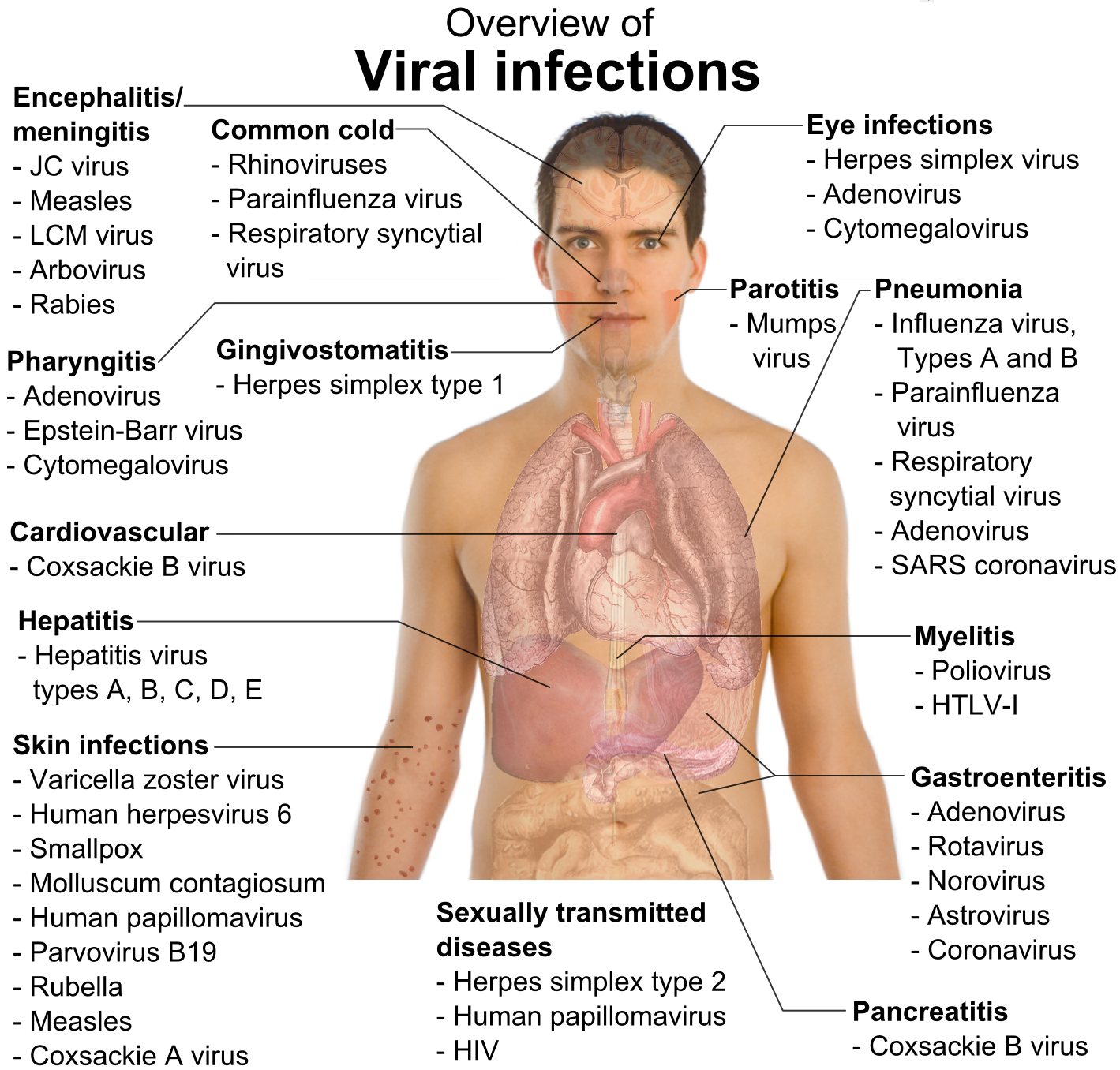
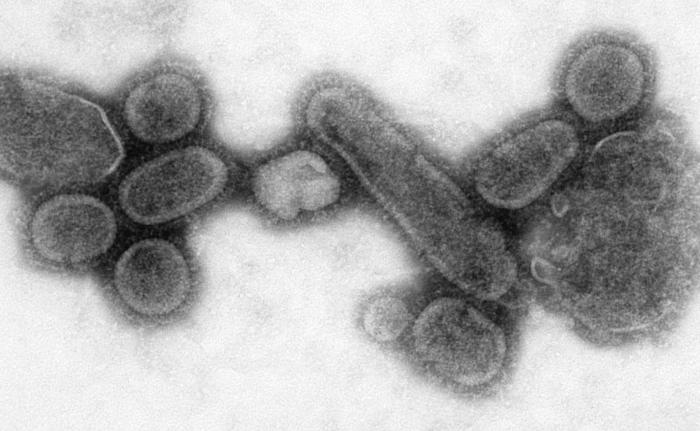


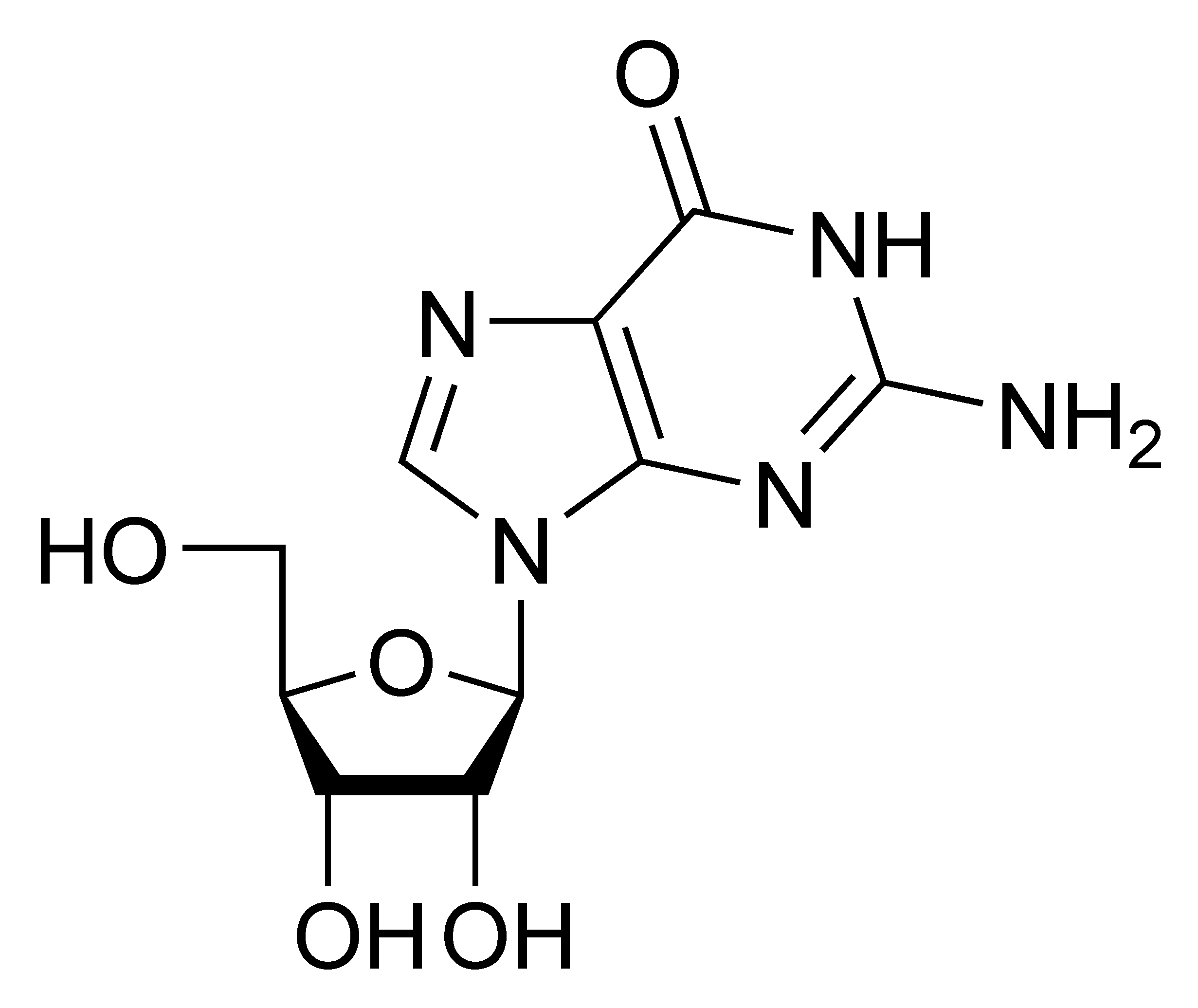

Wikimedia foundation. 2010.