- Tracheobionta
-
?Plantas vasculares (traqueófitas)

Clasificación científica Reino: Plantae Clados superiores: Archaeplastida
Viridiplantae
Streptophyta
Streptophytina
Embryophyta
Tracheobionta (Plantas vasculares)Divisiones - Rhyniophyta †
- Trimerophytophyta †
- "Pteridophyta"
- Lycophyta
- Euphyllophyta
- Monilophyta
- Psilotopsida
- Equisetopsida
- Marattiopsida
- Polypodiopsida
- Spermatophyta
- Gymnospermae teniendo en cuenta sólo los grupos vivientes (sensu stricto) es monofilético
- Pinophyta (coníferas)
- Cycadophyta
- Ginkgophyta
- Gnetophyta
- Magnoliophyta (Angiospermas)
- Gymnospermae teniendo en cuenta sólo los grupos vivientes (sensu stricto) es monofilético
- Monilophyta
Tracheobionta es el nombre del taxón de plantas que abarca a las traqueófitas, también llamadas plantas vasculares. Son organismos formados por células vegetales, que poseen un ciclo de vida en el que se alternan las generaciones gametofítica y esporofítica, siendo esta última la fase dominante (sobre quien actúa más presión de selección natural); cuya fase esporofítica es fotosintética e independiente, y tiene tejidos y sistemas de órganos; está organizada en un "cormo" (sistema que posee vástago aéreo, raíz subterránea y un sistema de conducción vascular que los vincula) que es a lo que comúnmente se refiere la gente cuando dice "planta"; cuya fase gametofítica es reducida y puede ser desde un "talo" (cuerpo no organizado en tejidos ni órganos) en helechos y afines, hasta unas pocas células protegidas y nutridas por el esporófito, en gimnospermas y angiospermas. La selección natural dirigió fuertemente la evolución de las traqueófitas hacia una menor dependencia de las condiciones ambientales sobre la tierra para la reproducción y la dispersión, característica que entra en evidencia al comparar las traqueofitas más antiguas (Lycophyta) con las más modernas (plantas con flores).
A lo largo de la historia se ha designado a este grupo con distintos nombres, como cormófitas o cormofitas (nombre científico Cormophyta o Cormobionta), embriófitas sifonógamas (nombre científico Embryophyta sifonogama), etc. Su nombre más común, Tracheophyta, proviene de tracheo (en referencia a las traqueidas, especializadas para el transporte de líquidos dentro de la planta) y phyta, raíz de origen griego que significa "planta". Se lo castellaniza como traqueofitas o traqueófitas.
La evidencia de los análisis moleculares de ADN hoy en día demostró que las traqueófitas son un grupo monofilético (que comprende a todos los descendientes de un ancestro común) dentro de las embriófitas, mientras que las briofitas son parafiléticas con respecto a las traqueofitas. Esto quiere decir que probablemente las traqueófitas sean descendientes de plantas muy parecidas a los briófitos, con el gametofito siendo la fase dominante, y el esporofito sin ramificar y dependiente nutricionalmente del gametofito.
Dentro de las traqueófitas hay dos linajes principales, Lycophyta y Euphyllophyta, diferenciadas principalmente por la forma de construcción de sus hojas (en las licofitas son lycofilos y en las eufilofitas son eufilos, los eufilos se corresponden aproximadamente con los megafilos, aunque en algunos grupos pueden estar reducidos en forma secundaria a su adquisición). Las eufilofitas a su vez comprenden dos grandes linajes, Monilophyta (helechos, equisetáceas y psilotáceas) y Spermatophyta, que se diferencian entre sí porque el primero tiene gametofitos de vida libre y el segundo los tiene encerrados en la semilla y el grano de polen. A su vez las espermatofitas están formadas por dos grandes grupos monofiléticos vivientes, Gymnospermae y Angiospermae o Magnoliophyta, que se diferencian entre sí porque el primer linaje tiene las semillas a la vista sobre la hoja fértil, mientras que el segundo tiene las semillas encerradas dentro de las paredes de la hoja fértil o carpelo.
Aún se siguen estudiando a las Lycophyta y Monilophyta como agrupadas en el grupo parafilético de los "helechos y afines" o Pteridophyta.
Las traqueofitas son un grupo importante tanto por dominar la mayor parte de los ecosistemas terrestres como por ser muy utilizadas por el hombre.
Contenido
El esporofito de las plantas vasculares
El cuerpo vegetativo
El cuerpo vegetativo del esporófito es un "cormo" (vástago + raíz + sistema de conducción vascular, hecho gracias al engrosamiento de las paredes celulares de células alargadas), que además de los tejidos vasculares posee tejidos de protección y tejidos de sostén, y que crece gracias a la acción de sus meristemas anatomy/11.html anatomy/14.html anatomy/13.html.
El "cormo" está especializado para la vida terrestre. Consta de vástago, raíz y un sistema de conducción vascular que los vincula. El vástago es la región del cormo que se especializa en la fotosíntesis, y que utiliza para ello el agua y las sales minerales que le llegan de la raíz. La raíz es la región del cormo que se especializa en la absorción de agua y sales, y que utiliza para ello los azúcares proporcionados por el vástago. Esta especialización es posible gracias a que el cormo posee un sistema de conducción vascular formado gracias a que las paredes de la célula vegetal pueden volverse ricas en lignina, compuesto que les otorga dureza.
Meristemas
El cormo crece gracias a la actividad de sus meristemas (del griego meros: dividir) que son un grupo de células en estado embrionario permanente, capaces de dividirse indefinidamente, formando tejidos que en su juventud son indiferenciados anatomy/11.html anatomy/14.html anatomy/13.html .
Los principales tipos de meristemas son dos:
- en todos los cormos encontraremos en los ápices de los tallos y las raíces, meristemas primarios responsables del crecimiento primario del esporofito.
- en los cormos con crecimiento secundario del tallo, encontraremos meristemas secundarios a lo largo del tallo, encargados de su crecimiento en grosor.
Sistemas de tejidos
El botánico J. Sachs en el siglo XIX, distinguió tres sistemas principales de tejidos en el cormo del esporofito de las cormófitas, clasificados según su función en la planta: tejidos de protección, tejidos fundamentales, y tejidos vasculares.
Tejidos de protección
Los tejidos que cumplen la función de protección forman la capa más externa del cormo.
- Si el cormo sólo posee crecimiento primario, encontraremos una epidermis, cubierta por una capa de cutina (lípido complejo que evita la pérdida de agua en la vida terrestre pero también evita el intercambio gaseoso con el medio ambiente), y con estomas Allies/Psilophyta/Psilotum nudum/Stem xs/Epidermis stomata.html y lenticelas (ambos aseguran el intercambio gaseoso con el ambiente).
- En los taxones con crecimiento secundario del tallo, la epidermis con cutina es reemplazada durante el crecimiento secundario, por una peridermis parcialmente impermeabilizada con suberina (lípido muy parecido a la cutina, responsable de la formación del súber o corcho).
Tejidos fundamentales
Los tejidos fundamentales forman un sistema continuo y están conformados principalmente por los diversos tipos de parénquimas (del griego: "carne de las vísceras"), a los que se asocian los tejidos de sostén (como colénquima y esclerénquima).
Los tejidos de sostén, como su nombre lo indica, cumplen la función de mantener la estructura de la planta, función que cumplen gracias a la lignina presente en la pared celular. Los tejidos de sostén más comunes son:
- Colénquima (del griego: goma, cola, nombre dado por la facilidad con que las paredes celulares se hinchan al hidratarse) formado por células vivas.
- Esclerénquima (del griego escleros: duro, nombre dado por sus gruesas paredes muy duras y resistentes) formado por células casi siempre muertas a su madurez.
Tejidos vasculares
Son los responsables del transporte de líquidos y sustancias por todo el cuerpo del vegetal, en el que se distinguen el floema (del griego floeos: "yema de la corteza, propio de la corteza", especializado en transporte de azúcares) y el xilema (del griego xylos: "lignificarse", formado por células muertas tubulares, abiertas o cerradas en sus extremos, de paredes muy lignificadas, que forman un haz vascular especializado en transporte de agua y sales) [1] [2] [3] [4] [5]. Estos tejidos son complejos, y a menudo, están asociados a otros (parenquimáticos y de sostén). Los tejidos vasculares se ubican dentro de los tejidos fundamentales de manera diversa según los distintos órganos de la planta (raíz, tallo, etcétera), que en un corte transversal forman patrones (llamados estela), que tienen importancia sistemática. Así el tallo de la mayor parte de las monocotiledóneas presenta un patrón o estela llamado atactostela, el tallo (primario) de las dicotiledóneas y de las coníferas en cambio, presenta una eustela, y el de los helechos y afines presenta una gran diversidad que permite diferenciar a las familias entre sí.
La reproducción en las plantas vasculares
 Diagrama esquemático del ciclo de vida de las plantas vasculares (Tracheobionta).
Diagrama esquemático del ciclo de vida de las plantas vasculares (Tracheobionta).
Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundaciónEl esporofito adulto desarrolla los llamados esporangios (estructuras multicelulares del esporofito dentro de las cuales se forman las esporas), donde se producirá la meiosis que dará esporas haploides. Las esporas al dividirse por mitosis se convierten en "gametofitos" (que serán un talo o sólo unas pocas células) que darán por meiosis las gametas masculina y femenina, la gameta femenina siempre será inmóvil y protegida al menos por el gametofito femenino dando en conjunto el óvulo, la gameta masculina será móvil o será transportada por agentes externos hasta el óvulo donde ocurre la fecundación, que dará un cigoto diploide. El nuevo esporofito se dividirá nutrido al principio al menos por el gametofito femenino, hasta dar el embrión formado por plúmula (donde está el meristema apical) y radícula (donde está el meristema radical), que al crecer forma el nuevo esporofito adulto recomenzando el ciclo. En los taxones sin hojas (Psilotum) los esporangios se ubican directamente sobre el tallo, en los taxones con hojas, los esporangios se ubican en las hojas (convirtiéndose éstas en "esporofilos").
Dentro de los esporangios se produce la meiosis que forma las esporas haploides, que luego serán liberadas o no, y al dividirse por mitosis darán gametofitos haploides. En las cormófitas más antiguas (como los helechos y afines) la fase gametofítica todavía es de vida libre, pero en las más recientes se ha vuelto totalmente dependiente del cuerpo vegetativo del esporofito para su nutrición y protección. Las pteridofitas (helechos y afines) pueden ser isospóricas (todas las esporas iguales, darán gametofitos hermafroditas) o heterospóricas (con dos morfos de esporas, megasporas que darán gametofito femenino y microesporas que darán gametofito masculino); en cambio las espermatofitas (gimnospermas y angiospermas) son todas heterospóricas. Cuando el esporofito es heterospórico, el gametofito se desarrolla por completo dentro de la espora, sin ser de vida libre. Los gametofitos producirán las gametas femeninas y masculinas (del griego gametes: marido). La gameta femenina inmóvil espera ser fecundada dentro de una estructura donde está protegida y será nutrida (fenómeno de oogamia), esa estructura proviene del gametofito (en pteridofitas) o del gametofito y del esporofito (en espermatofitas). Al recibir a la gameta masculina se produce la fecundación que dará el cigoto diploide de un nuevo esporofito (del griego zigos: pareja, yugo). El cigoto se empezará a dividir pasando por una fase de embrión antes de convertirse en adulto, el embrión se nutre de sus cubiertas protectoras. Sólo en las espermatofitas el embrión puede mantenerse en estado de latencia (en forma de "semilla") hasta que las condiciones sean adecuadas (y se produzca lo que se conoce como "germinación"), las pteridofitas, más primitivas, se desarrollan directamente hasta adultas sin latencia. En el embrión ya se distinguen los meristemas primarios: el meristema apical que da origen al vástago que también da origen a las hojas si las hay anatomy/15.html, y un meristema radical que da origen a la raíz primaria anatomy/19.html. La raíz primaria o raíz embrionaria puede desarrollarse o morir, ya que el vástago también puede emitir raíces que en este caso se llamarán raíces adventicias.
Sistemática
Las cormófitas pueden dividirse en dos grandes grupos según el sistema Engler:
- Grupo parafilético Pteridophyta. La hipótesis más aceptada hasta hace unos años sostenía que las pteridofitas son un grupo monofilético derivado de algún antecesor del alga verde. Hoy en día se sabe compuesta por dos líneas filogenéticas, son las más antiguas de las cormófitas actuales, comprende a los helechos y afines. Se caracterizan por poseer una alternancia de generaciones bien manifiesta. Las pteridofitas actuales están concentradas en los trópicos y áreas montanas húmedas. Está compuesta por las divisiones monofiléticas:
- División Lycopodiophyta
- División Monilophyta
- Superdivisión Spermatophyta. Las espermatofitas o antofitas pueden definirse como cormófitas donde la alternancia de generaciones se da en forma enmascarada, ya que el gametofito se desarrolla dentro de las estructuras del esporofito (incluso en los grupos más primitivos todavía pueden observarse anteridios y arquegonios). Comprende a las gimnospermas y angiospermas, en las siguientes líneas monofiléticas:
- División Gymnospermae
- División Magnoliophyta (angiospermas)
Los últimos análisis genéticos determinaron que la fiolegnia de las traqueófitas es la siguiente:
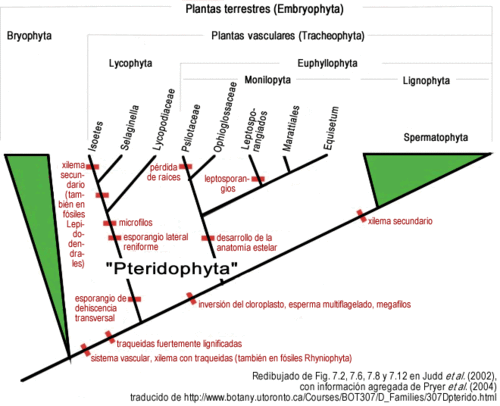
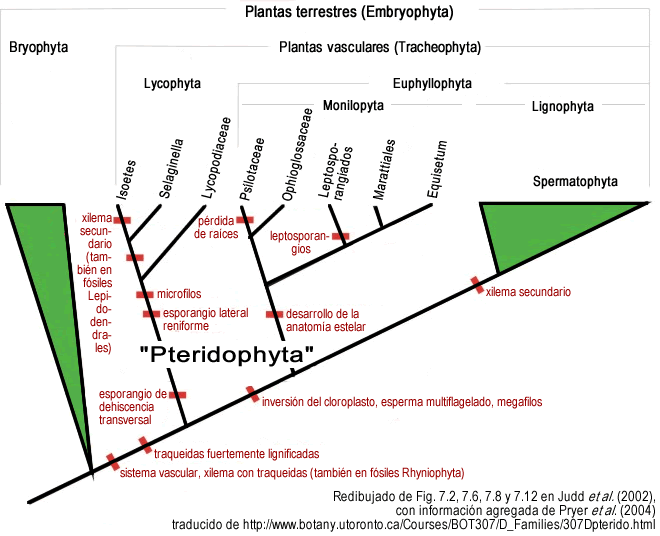
Figura 5. Árbol filogenético actualizado de los diferentes grupos de pteridofitos y su relación con los demás grupos de plantas terrestres. En particular es de destacar que una evidencia molecular muy fuerte de la división basal entre licofitas y eufilofitas es la presencia en las eufilofitas de una inversión de unas 30 kilobases en el ADN del cloroplasto (Raubeson y Jansen 1992), lo notable es que inversiones de tantas kilobases son muy raras, y es raro que den productos viables, por lo que se convierte en evidencia muy fuerte de monofilia.
Evolución de traqueófitas
 Reconstrucción del esporofito de Rhynia. Aspecto general, corte longitudinal del esporangio, tétrada de esporas y detalle de espora con marca trilete. Nótense la ramificación dicotómica del tallo, la falta de hojas y raíces, y los esporangios terminales.
Reconstrucción del esporofito de Rhynia. Aspecto general, corte longitudinal del esporangio, tétrada de esporas y detalle de espora con marca trilete. Nótense la ramificación dicotómica del tallo, la falta de hojas y raíces, y los esporangios terminales.Mucho de lo que sabemos acerca de las primeras plantas vasculares y su transición a la vida terrestre se encuentra en el registro fósil, ya que esos valiosos linajes están todos extintos.
Las traqueófitas ancestrales derivaron de las primeras embriófitas, las primeras plantas que colonizaron la superficie terrestre. Todo indica que las primeras embriófitas eran pequeñas y muy simples en su estructura. En el caso del linaje que derivó en las plantas vasculares, el esporofito era básicamente un tallo de ramificación dicotómica, al principio de la altura de un fósforo, con el esporangio (donde ocurre la meiosis que da esporas haploides) producido en las puntas de las ramas. Estas plantas no tenían ni hojas ni raíces, en algunos casos (como Rhynia del "Rhynia Chert" en Escocia) la preservación de estas plantas es espectacular, y es posible discernir muchos detalles anatómicos, incluyendo estomas, esporas, y el tejido vascular dentro del tallo.
En los análisis basados en dichos fósiles, fue recientemente descubierto que los primeros polysporangiofitos (plantas con esporofitos ramificados) en realidad no producían verdaderas células conductoras de agua (traqueidas) en el xilema, y por lo tanto deben haber dependido por completo de la presión de turgencia para mantenerse erguidos. Las células verdaderamente conductoras de agua evolucionaron más tarde y caracterizan al clado de las traqueófitas o "verdaderas plantas vasculares" (Kenrick y Crane 1997a,[1] 1997b[2] ).
Las traqueidas son células elongadas con paredes engrosadas, muertas a la madurez. Donde una traqueida se conecta con la siguiente se observan unas aberturas características en la pared secundaria ("pits" en inglés) pero la pared celular primaria de las células que formaron las traqueidas se mantiene intacta, dando la "pit membrane". El agua por lo tanto debe atravesar las paredes primarias de las células para poder avanzar. En las primeras traqueófitas (representadas por Rhynia)las traqueidas era de un tipo distintivo, con cierta "resistencia al desmoronamiento" ("decay resistance"), conferida por la lignificación de las fibras de celulosa, que estaba presente sólo como una capa muy delgada. Las paredes celulares más resistentes son las que caracterizan al clado de las eutracheofitas, que include a todas las plantas vasculares vivientes (Kenrick y Crane 1997a[1] ). En estas especies, las traqueidas fuertemente lignificadas permiten una conducción de agua más eficiente, y proveen resistencia interna, permitiendo a las plantas crecer mucho más altas.
Para leer una discusión acerca de la evolución de la predominancia del esporofito en detrimento del gametofito, ver Evolución de embriófitas.
Las relaciones filogenéticas entre las líneas de plantas vasculares vivientes, mostradas en la figura de la sección sistemática, muestran una división basal, que ocurrió en el Devónico temprano a medio (hace unos 400 millones de años), que separa al linaje de las lycophytas actuales del linaje de las eufilofitas actuales. Esta división está marcada por una considerable variedad de características morfológicas. Una bastante notoria es la presencia de esperma multiflagelado en las eufilofitas en oposición al esperma biflagelado de las lycophytas y de los linajes desprendidos de grupos más antiguos (los briófitos sensu lato). Las dos únicas excepciones son Isoetes y Phylloglossum, donde el esperma multiflagelado se originó independientemente.
Las primeras plantas con semilla aparecieron en el Devónico tardío, y eso llevó a la típica flora del Mesozoico dominada por gimnospermas. Los fósiles más antiguos de angiospermas encontrados hasta la fecha datan aproximadamente de hace 140 millones de años (principios del Cretácico), y el número de especies encontrado aumenta de forma llamativa con el tiempo geológico, en comparación a otras plantas no angiospermas (el llamado "abominable misterio" de Darwin). Para una discusión sobre la radiación de las angiospermas ver Evolución de angiospermas.
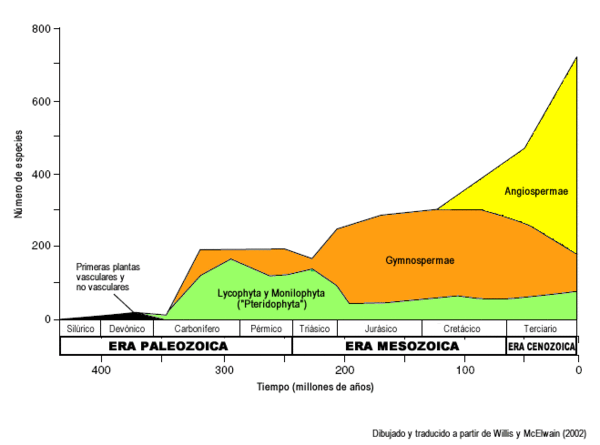
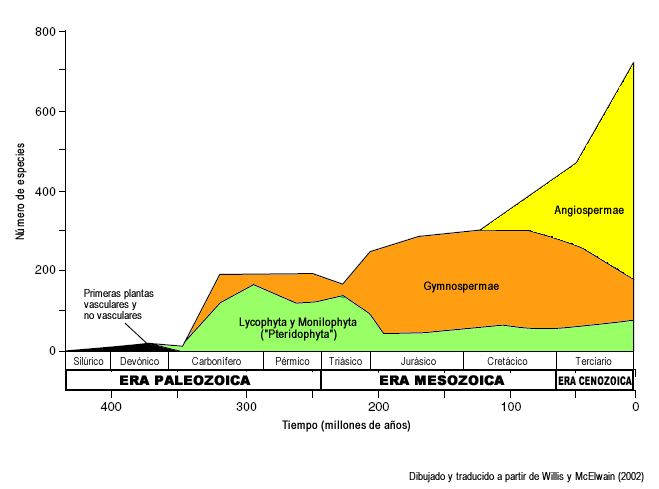
Registro fósil de aparición de los diferentes clados de plantas vasculares. Nótese la intensa radiación de las angiospermas en comparación con los helechos y gimnospermas. Dibujado y traducido a partir de Willis y McElwain (2002).[3] Importancia económica
Las traqueofitas son un grupo importante para el hombre, además de dominar la mayor parte de los ecosistemas terrestres: nos provee de medicinas, plantas ornamentales, fibras para papel y ropa, y la mayor parte de nuestros alimentos. Históricamente, la información sobre éstos y otros atributos de las traqueofitas fue esencial para el desarrollo de la civilización humana. La supervivencia dependía de conocer cuáles eran las plantas que eran buenas para comer, cuáles eran venenosas para la gente o para los animales que eran potencial alimento, cuáles eran buenas para armas y herramientas, cuáles podían curar, y cuáles podían ser útiles en muchos otros sentidos.
Enlaces externos
- P. F. Stevens, 2001 en adelante, Angiosperm Phylogeny Website. Versión 9, junio del 2008, y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/
Bibliografía
- Judd, W. S.; Campbell, C. S.; Kellogg, E. A.; Stevens, P. F.; Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- H. C. Bold, C.J. Alexopoulos y Th. Delevoryas (1989) Morfología de las Plantas y los Hongos. Omega, Barcelona, 911 pp., ISBN 84-282-0754-2
- Juan J. Valla, BOTÁNICA, Morfología de las Plantas Superiores, editorial Hemisferio Sur, Buenos Aires, Argentina, 1979.
- 2000. The PLANTS Database, database (version 5.1.1). National Plant Data Center, NRCS, USDA. Baton Rouge, LA 70874-4490 USA. http://plants.usda.gov
- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (resumen en inglés aquí).
Referencias citadas
- ↑ a b Kenrick, P y PR Crane. 1997a. The origin and early diversification of land plants: A cladistic study. Smithsonian Institution Press, Washington, DC.
- ↑ Kenrick, P y PR Crane. 1997b. The origin and early evolution of plants on land. Nature 389: 33-39.
- ↑ Willis y McElwain. 2002. "Flowering Plant Origins". en: The Evolution of Plants, capítulo 6.
Caracteristicas generales de las traqueofitas inferiores y superiores

Wikimedia foundation. 2010.