- Hipótesis del mundo de ARN
-
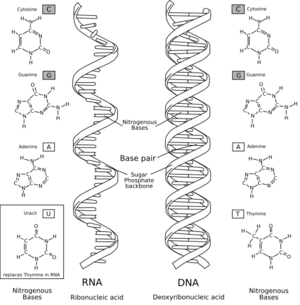 ARN con sus bases nitrogenadas a la izquierda y ADN a la derecha.
ARN con sus bases nitrogenadas a la izquierda y ADN a la derecha.
La hipótesis del mundo de ARN propone que el ARN fue la primera forma de vida en la Tierra, desarrollando posteriormente una membrana celular a su alrededor y convirtiéndose así en la primera célula procariota.
Contenido
Historia
La expresión "Mundo de ARN" fue empleada por primera vez por el premio nobel Walter Gilbert en 1986, al hilo de un comentario sobre las recientes observaciones sobre las propiedades catalíticas de algunas formas de ARN.[1] No obstante, la idea de una vida independiente basada en el ARN es más antigua y ya se puede encontrar en El código genético, de Carl Woese.[2] En 1963 el biólogo molecular Alexander Rich, del MIT especuló sobre la misma idea en un artículo en el que contribuía a un volumen publicado en honor al fisiólogo y premio Nobel Albert Szent-Györgyi.
Propiedades del ARN
Las propiedades del ARN nos pueden dar una idea de la posibilidad conceptual de la hipótesis del mundo de ARN, aunque su plausibilidad como explicación del origen de la vida se encuentra debatida. Se sabe que el ARN es un eficiente catalizador y al igual que el ADN posee la capacidad de almacenar información.
Una versión de la hipótesis ligeramente distinta es que un tipo diferente de ácido nucleico, denominado "pre-ARN" fuera el primero en surgir como molécula autorreproductora para ser reemplazado por el ARN sólo después. Estos ácidos nucleicos a menudo se producen y polimerizan más fácilmente bajo condiciones prebióticas. Algunos candidatos serían el ANP, el ANT y el ANG.[3] [4]
El ARN como enzima
Las enzimas de ARN o ribozimas son posibles en el actual mundo de ADN, aunque no comunes. No obstante las ribozimas desempeñan papeles de vital importancia. Las ribozimas son componentes esenciales del ribosoma, el cual es esencial para la síntesis de proteínas. Son posibles muchas funciones de los ribosomas: La naturaleza emplea ampliamente el auto-splicing del ARN y la evolución dirigida ha creado ribozimas con una gran variedad de actividades. Entre las propiedades enzimáticas importantes para el comienzo de la vida se encuentran:
- La capacidad de la autoduplicación o de duplicar otras moléculas de ARN. Se han producido en el laboratorio moléculas relativamente cortas de ARN. Las más cortas eran de una longitud de 165 bases aunque se ha estimado que sólo parte de estas bases eran cruciales para esta función. Una versión cuya longitud era de 189 pares de bases obtenía una fidelidad de copia del 98.9%,[5] lo que significaría que podría hacer una copia exacta de una molécula de ARN tan larga como ella misma de cada ocho copias, aunque esta ribozima de 189 bp como mucho podría polimerizar un molde de 14 nucleótidos de longitud, demasiado corto para considerarlo replicación, pero representa un gran comienzo. La mayor extensión de un cebador llevada a cabo por una ribozima de tipo polimerasa fue de 20 bp.[6]
- La capacidad de catalizar reacciones químicas sencillas que aumentan la creación de los monómeros del ARN. Se han creado en el laboratorio moléculas relativamente pequeñas de ARN en el laboratorio con esa capacidad de manera artificial.[7] [8]
- La capacidad de formar enlaces peptídicos para producir péptidos cortos y ocasionalmente proteínas enteras. Esto se efectúa en las células modernas por los ribosomas, un complejo de dos moléculas grandes de ARN conocidas como ARN ribosómico junto con otras proteínas. Se piensa que las dos moléculas de ARNr son las responsables de esta actividad enzimática. Se ha creado en el laboratorio una molécula de ARN más corta capaz de formar enlaces peptídicos, y se ha sugerido que el rARN evolucionó a partir de una molécula similar.[9] Así, también se ha sugerido que los aminoácidos podrían inicialmente complementarse con moléculas de ARN como cofactores mejorando o diversificando sus capacidades enzimáticas, antes de evolucionar a péptidos más complejos; el RNAm puede haber evolucionado con a partir de tales moléculas de ADN, y el ARNt de moléculas que hayan catalizado transferencias de aminoacidos de ellos.[10]
ARN como almacén de información
El ARN es una molécula muy parecida al ADN y guarda dos diferencias químicas con éste: la estructura completa del ARN y del ADN son inmensamente similares. Una cadena de ADN y otra de ARN pueden unirse para formar una estructura de doble hélice. Esto hace posible el almacenamiento de información en el ARN de una forma muy parecida a la que se efectúa en el ADN.
Comparaciones de las estructuras del ADN y ARN
La principal diferencia es la presencia de un grupo hidroxilo en posición 2' del azúcar ribosa en el ARN. Este grupo hace que la molécula sea menos estable. En las regiones flexibles del ARN (por ejemplo, las que no están constreñidas por la doble hélice) pueden ser atacadas químicamente por el enlace fosfodiéster adyacente de modo que se puede escindir. El grupo hidroxilo también fuerza a la ribosa a adoptar la conformación C3'-endo en lugar de la más habitual C2'-endo del ADN. Esto fuerza a la doble hélice a adoptar una conformación ligeramente diferente a la del ADN.
El ARN también utiliza un grupo de bases diferente -adenina, guanina, citosina y uracilo- en lugar de adenina, guanina citosina y timina. El uracilo es químicamente similar a la timina, aunque emplea menos energía para producirse. En términos del apareamiento de bases esto no tiene ningún efecto, ya que la adenina puede unirse al uracilo perfectamente. Sin embargo, el uracilo es un producto de la trasmutación por daños de la citosina, haciendo que el ARN sea particularmente susceptible a mutaciones que pueden reemplazar un par de bases GC por GU o AU.
Limitaciones en el almacenamiento de información en el ARN
No es fácil almacenar grandes cantidades de información en el ARN. Las propiedas químicas del ARN hacen que las moléculas grandes de ARN sean inherentemente frágiles y se puedan fragmentar con facilidad, siendo posteriormente descompuestas en nucleótidos por hidrólisis. Las bases aromáticas también absorben fuertemente la fracción ultravioleta del espectro y podrían haber sido susceptibles de daños y descomposición por la radiación de fondo.[11] [12] Estas limitaciones no hacen que sea imposible la utilización del ARN para almacenar información, sino solamente exigente desde el punto de vista energético (para reparar o reemplazar las moléculas dañadas de ARN) y propensas a la mutación. Mientras que esto hace que sea poco adecuado en la actual vida "optimizada" basada en el ADN, lo pudo haber sido para la vida primitiva.
Pruebas
La hipótesis del mundo de ARN se basa en la capacidad del ARN de almacenar, transmitir y duplicar la información genética, de la misma forma que lo hace el ADN. El ARN puede actuar también como una ribozima (una enzima hecha de ácido ribonucleico). Debido a que puede reproducirse a sí misma, desarrollando las tareas del ADN y de las proteínas (enzimas), se piensa que el ARN fue capaz de tener su propia vida independiente. Así, mientras que no se encontraron nucleótidos en el Experimento de Miller y Urey, sí se encontraron en las simulaciones de otros investigadores, sobre todo en las de Juan Oró. Experimentos con los ribozimas básicos, como el ARN viral Q-beta, han demostrado que las estructuras de ARN autorreplicantes sencillas pueden resistir incluso a fuertes presiones selectivas (como los terminadores de cadena de quiralidad opuesta).[13]
Walter Gilbert fue el primero en usar la expresión "mundo de ARN" en 1986. No obstante, la idea de vida ARN independiente es más antigua, y se puede encontrar en el libro de Carl Woese The Genetic Code.[14] Cinco años antes, el biólogo molecular Alexander Rich, del Massachusetts Institute of Technology, ya había propuesto gran parte de la misma idea en un artículo escrito para un volumen publicado en honor del premio Nobel de Fisiología Albert Szent-Györgyi.
Dificultades con la hipótesis
Puesto que no existen rutas químicas para la síntesis abiogénica de nucleótidos a partir de las bases pirimidínicas de citosina y uracilo bajo condiciones prebióticas podría ser que los ácidos nucleicos no las contuvieran.[15] Resulta revelador que el nucleósido citosina tenga una vida media en aislamiento de 19 días a 100 °C y 17.000 años en agua helada, lo cual es aún muy corto para los tiempos a escala geológica.[16] Otros autores ponen en cuestión que la ribosa y otros azúcares del esqueleto de los ácidos nucleicos puedan ser lo suficientemente estables como para encontrarse entre el material genético original.[17] Por ejemplo, se sabe que el enlace éster entre la ribosa y el ácido fosfórico en el ARN es propenso a la hidrólisis.[18] Además, la ribosa tiene que ser toda del mismo enantiómero, porque cualquier nucleótido con una quiralidad diferente actuaría como terminador de la cadena.[19]
Detalles del mundo de ARN
Mecanismos propuestos para la síntesis prebiótica de ARN
Los nucleótidos son moléculas fundamentales que se combinan en serie para formar ARN. Consisten en una base nitrogenada unida a un esqueleto de azúcar-fosfato. El ARN está compuesto por largos tramos de nucleótidos específicos ordenados de tal modo que su secuencia de bases porta información. La hipótesis del mundo de ARN sostiene que en el caldo primigenio o sandwich primigenio existirían ya los nucleótidos en disolución. Estos nucleótidos formarían enlaces de forma regular con otros, que se romperían porque el cambio de energía era bajo. No obstante, ciertas secuencias de pares de bases tendrían propiedades catalíticas que disminuirían la energía para generar su cadena, haciendo que permanecieran juntos por periodos de tiempo más largos. A medida que se alargaba la cadena iba atrayendo los nucleótidos que encajaban de forma más rápida, haciendo que la cadena se formara más rápidamente que su velocidad de degradación.
Se ha propuesto que estas cadenas eran las primeras y más primitivas formas de vida. En el mundo de ARN las diferentes cadenas competían entre ellas por los nucleótidos libres y estaban sujetas a la selección natural. Las moléculas más eficientes de ARN, aquellas capaces de catalizar eficientemente su propia reproducción, sobrevivían y evolucionaban, formando el moderno ARN.
La competición entre los ARN pudo haber favorecido el surgimiento de una cooperación entre cadenas diferentes, abriendo la vía para la formación de las primeras protocélulas. En algún momento las cadenas de ARN desarrollaron al azar propiedades catalíticas que ayudaban a los aminoácidos a unirse mediante enlaces peptídicos. Estos aminoácidos podrían entonces contribuir a la síntesis de ARN, proporcionando a esas cadenas de ARN que actuaban como ribozimas una ventaja selectiva. Con el tiempo fueron reclutadas para la vida el ADN, los lípidos, los carbohidratos y todos los tipos de sustancias químicas que hoy forman parte de ella. Esto condujo a la aparición de las primeras células procariotas y finalmente a la vida tal y como la vemos actualmente.
Desarrollos posteriores de la hipótesis
Patrick Forterre ha estado trabajando en una hipótesis controvertida que establece que los virus fueron el instrumento que produjo la transición del ARN al ADN y la evolución de las Bacterias, Archaeas y Eucariotas. Este autor cree que el último antepasado común fue un ser basado en el ARN que desarrolló virus de ARN. Alguno de estos virus evolucionaron en virus de ADN para proteger sus genes de ataques. A través del proceso de infección viral evolucionaron los tres dominios de la vida.[20]
Hipótesis alternativa
Como se ha mencionado anteriormente, una versión distinta de la misma teoría es la del "mundo de pre-ARN", en la que se propone que un ácido nucleico diferente precedió al ARN. Como alternativa se ha propuesto al Ácido nucleio peptídico. El ANP es más estable que el ARN y parece que sería más sencillo de sintetizar en condiciones prebióticas, especialmente porque la síntesis de ribosa y la adición de grupos fosfatos son problemáticas, porque no existían ninguna de las dos. Se ha propuesto el Ácido nucleico de Treosa como punto de partida, así como el ácido nucleico de glicerol.
Se ha propuesto una alternativa diferente -o complementaria- al ensamblaje de ARN en la Hipótesis del mundo de HAP. Otra teoría alternativa más a la hipotesis del ARN sería la hipótesis de la panspermia. Explora la posibilidad de que el origen de la vida en este planeta fuera traída de algún otro lugar de la galaxia.
Implicaciones del mundo de ARN
De ser cierta la hipótesis del mundo de ARN tendría importantes implicaciones para la misma definición de la vida. Durante la mayor parte del tiempo que siguió al descubrimiento de la estructura del ADN por Watson y Crick, se consideró que la vida se definía en gran medida en términos de ADN y proteínas. El ADN y las proteínas parecián ser las macromoléculas dominantes en las células vivas, de modo que el ARN tenía el único propósito de ayudar a crear proteínas a partir del "plano" de ADN. Esta hipótesis sitúa al ARN en el papel central en el origen de la vida, lo cual ha promovido numerosos estudios en los últimos diez años que han desvelado importantes aspectos en la función del ARN que previamente se desconocían y demuestran un papel crítico del ARN en la funcionalidad de la vida. En 2001 la hipótesis del mundo de ARN recibió un gran impulso al descifrarse la estructura tridimensional del ribosoma, que revelaba que los lugares catalíticos claves del ribosoma estaban hechos de kARN y que las proteínas no tenían un papel estructural importante y eran de importancia funcional accesoria. Específicamente hablando, se sabe actualmente que la formación del enlace peptídico, que es la reacción que une los aminoácidos entre sí para formar proteínas está catalizada por un residuo adenina del ARN ribosómico y por tanto el ribosoma es una ribozima. Este descubrimiento sugiere que las moléculas de ARN fueron con toda probabilidad capaces de generar las primeras proteínas. Otros descubrimientos importantes mostraron que el ARN era más que un simple mensajero o adaptador para transferir moléculas, como es en el caso de las riboproteínas nucleares pequeñas (snRNP) en el procesamiento del pre-mARN y la corrección del ARN (ARN editing) y la transcripción del ARN en Eucariotas en el mantenimiento de los telómeros y en la reacción de la telomerasa.
Véase también
- Conjunto autocatalítico
- Origen de la vida
- Teoría de la generación espontánea
- Experimento de Miller y Urey
- Teoría del mundo de hierro-sulfuro
- Hipótesis del mundo de HAP
- Panspermia
Referencias
- ↑ Gilbert, Walter (Feb 1986). «The RNA World». Nature 319: pp. 618. doi:.
- ↑ Woese, Carl (Jan de 1968). The Genetic Code. Harper & Row. ISBN 978-0060471767.
- ↑ Orgel, Leslie (Nov 2000). «A Simpler Nucleic Acid». Science 290 (5495): pp. 1306-7. doi:.
- ↑ Nelson, K.E.; Levy, M.; Miller, S.L. (Apr 2000). «Peptide nucleic acids rather than RNA may have been the first genetic molecule». Proc. Natl. Acad. Sci. USA 97 (8): pp. 3868–71. PMID 10760258. http://www.pnas.org/cgi/content/abstract/97/8/3868.
- ↑ W. K. Johnston, P. J. Unrau, M. S. Lawrence, M. E. Glasner and D. P. Bartel RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension. Science 292, 1319 (2001)
- ↑ Hani S. Zaher and Peter J. Unrau, Selection of an improved RNA polymerase ribozyme with superior extension and fidelity. RNA (2007), 13:1017-1026
- ↑ Huang, Yang, and Yarus, RNA enzymes with two small-molecule substrates. Chemistry & Biology, Vol 5, 669-678, November 1998
- ↑ Unrau, P.J. and Bartel, D.P. (1998) RNA-catalysed nucleotide synthesis. Nature 395, 260-263
- ↑ Zhang and Cech, Peptide bond formation by in vitro selected ribozymes. Nature 390, 96-100
- ↑ Szathmary E., The origin of the genetic code: amino acids as cofactors in an RNA world. Trends in Genetics, Volume 15, Number 6, 1 June 1999 , pp. 223-229(7)
- ↑ Lindahl, T (Apr 1993). «Instability and decay of the primary structure of DNA». Nature 362 (6422): pp. 709-15. PMID 8469282.
- ↑ Plantilla:Citejournal
- ↑ The Basics of Selection (London: Springer, 1997)
- ↑ New York: Harper and Row, 1967
- ↑ L. Orgel, The origin of life on earth. Scientific American. 271 (4) p. 81, 1994.
- ↑ Matthew Levy and Stanley L. Miller, The stability of the RNA bases: Implications for the origin of life, Proceedings of the National Academy of Science USA 95, 7933–7938 (1998)
- ↑ Larralde R, Robertson M P, Miller S L. Proc Natl Acad Sci USA. 1995;92:8158–8160.
- ↑ Lindahl T. Nature (London). 1993;362:709–715.
- ↑ Joyce GF; Visser GM, van Boeckel CA, van Boom JH, Orgel LE, van Westrenen J. (Aug 1984). «Chiral selection in poly(C)-directed synthesis of oligo(G)». Nature 310 (5978): pp. 602-4. PMID 6462250.
- ↑ Zimmer C. (2006). «Did DNA come from viruses?». Science 312 (5775): pp. 870-2. PMID 16690855.
Bibliografía
- Cairns-Smith, A. G. Genetic Takeover: And the Mineral Origins of Life. ISBN 0-521-23312-7
- Lindahl, T., 1993. "Instability and decay of the primary structure of DNA," Nature 362(6422):709–715.
- Pääbo, S. 1993. "Ancient DNA" Scientific American 269(5):60–66.
Enlaces externos
- The RNA World Website
- "Self-replication: Even peptides do it" por Stuart A. Kauffman, en Nature 382 August 8, 1996
- "The RNA world" (2001) por Sidney Altman, en la página web de los Premios Nobel
- "Exploring the new RNA world" (2004) por Thomas R. Cech, en la página web de los Premios Nobel
- "The Beginnings of Life on Earth" (1995) por Christian de Duve en American Scientist Online
- "The Origin of Life on the Earth" por Leslie E. Orgel
- "The Formation of the RNA World" por James P. Ferris
Wikimedia foundation. 2010.