- Hipótesis fagotrófica
-
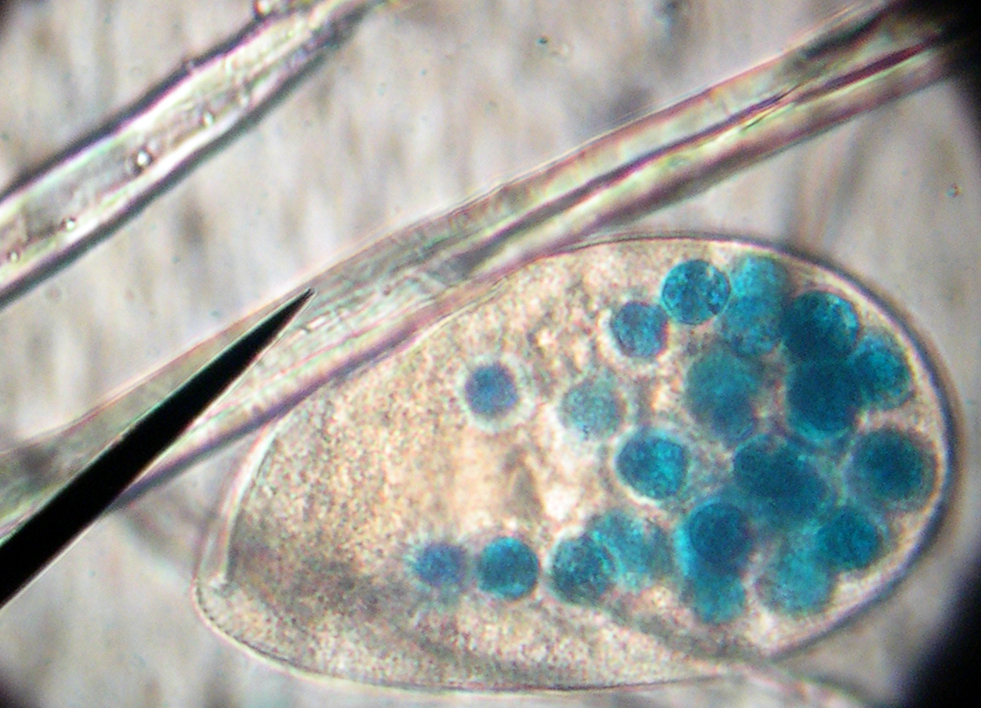
 Este paramecio alimentado con alimento tintado muestra sus vacuolas en color azul.
Este paramecio alimentado con alimento tintado muestra sus vacuolas en color azul.
La hipótesis fagotrófica postula que el origen de la célula eucariota fue conducido por la capacidad de ingerir presas.[1] Según esta hipótesis, la conversión de la rígida pared bacteriana de peptidoglicano en una superficie celular flexible de glicoproteínas permitió por primera vez en la historia de la vida la fagocitosis.
La capacidad de ingerir presas está unida a la endocitosis que llevó necesariamente al desarrollo del sistema de endomembranas, al citoesqueleto y a sus motores moleculares asociados, y en última instancia, dio lugar al núcleo y a la reproducción sexual. Es decir, que la mayor parte de la características eucariotas tendrían un origen autógeno. La fagotrofia sería también responsable de la adquisición de las mitocondrias y cloroplastos, pues abre inmediatamente la posibilidad de que alguna célula ingerida escape a la digestión y se convierta, bien en un parásito intracelular, bien en un endosimbionte.
Contenido
Origen
Según Cavalier-Smith,[1] las innovaciones radicales que dieron lugar a los eucariontes tuvieron como partida un antecesor Neomura, del cual se derivaron tanto Archaea como Eukarya, que serían, por tanto, grupos hermanos. Estas innovaciones incluyen la fagocitosis, sistema de endomembranas incluyendo peroxisomas, citoesqueleto, núcleo celular y reproducción sexual. Cavalier-Smith también supone que el antecesor neomura se derivaría de una actinobacteria. Por tanto, rechaza la idea de que los eucariontes se originaran por la fusión de una archaea y una proteobacteria alfa, como postula la hipótesis de endosimbiosis serie y que en su opinión falla en explicar las numerosas características compartidas específicamente entre eucariontes y actinobacterias.
El paso de Bacteria a Neomura vendría marcada por veinte adaptaciones evolutivas, consecuencia de otras dos adaptaciones importantes: el desarrollo de histonas para reemplazar la ADN girasa y la sustitución de la pared celular de peptidoglucano por otra de glicoproteína (neomura significa "nueva pared").[2] Pero las archaea se adaptaron a ambientes calientes y ácidos, reemplazando los lípidos acilo éster de las bacterias por lípidos prenil éter, y usaron las glicoproteínas como una nueva pared pared rígida, y por tanto, retuvieron la organización celular bacteriana. Los eucariontes, en cambio, usaron la nueva superficie de proteínas como una capa flexible que dio lugar a la fagocitosis y que llevó, en última instancia, al cambio en la estructura de la célula.
El antecesor más probable para neomura, y por lo tanto para los eucariontes, podría ser un derivado de una bacteria Gram-positiva heterótrofa y aerobia, más que un metanógeno anaerobio que necesitaría más cambios metabólicos. Una actinobacteria es el antecesor más probable, puesto que es la única bacteria que produce quitina, además su ácido graso sintetasa es un agregado macromolecular y no moléculas separadas como en el resto de las bacterias y la formación de exosporas puede interpretarse como un precursor de las zigosporas, probablemente el primitivo método de reproducción sexual de los eucariontes.
Las actinobacterias también sintetizan colesterol (al igual que otros dos grupos de proteobacterias), un compuesto complejo que es poco probable que haya surgido dos veces a lo largo de la evolución. Los eucariontes utilizan el colesterol en sus membranas, y es probable que sus antecesores dieran esta nueva función a este compuesto cuando desarrollaron la fagocitosis y las endomembranas. Además, esto también implica que los fósiles de hace 2.700 millones de años que contienen esteranos y que eran anteriormente considerados como un indicio de la presencia de eucariontes, podrían proceder de bacterias y el origen de los eucariontes ser mucho más reciente.
El siguiente árbol filogenético muestra la posición de Eukarya en la teoría de la evolución bacteriana de Cavalier-Smith.[3] [4]
[A] Chlorobacteria
[B] Hadobacteria
[C] [D] [E] [F] [G] [H] [I] Endobacteria
[J] [K] Neomura [L] [M] Eukarya
Leyendas: [A] Bacteria Gram-negativa con pared de peptidoglicano y clorosomas. [B] Fotosíntesis oxigénica, Omp85 y cuatro nuevas catalasas. [C] Revolución glicobacteriana: membrana externa con inserción de lipopolisacáridos, hopanoides, ácido diaminopimélico, ToIC y TonB. [D] Ficobilisomas. [E] Flagelos. [F] Cuatro insecciones: un aminoácido en Hsp60 y FtsZ y un dominio en las ARN polimerasas β y σ. [G] Endosporas. [H] Bacterias Gram-positivas: hipertrofia de la pared de peptidoglicano, sortasas y pérdida de la membrana externa. [I] Glicerol 1-P deshidrogenasa [J] Proteasomas y fosfatidilinositol. [K] Revolución Neomura: peptidoglicano y lipoproteínas son sustituidos por glicoproteínas. [L] ADN girasa inversa y lípidos éter isoprenoides. [M] Fagotrofia.
Transformación de la pared celular, citoesqueleto y motores moleculares
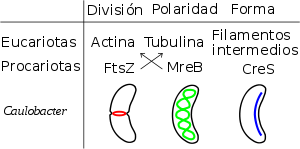 Elementos del citoesqueleto de Caulobacter crescentus. En la figura, estos elementos procarióticos se relacionan con sus homólogos eucariotas y se hipotetiza su función celular.[5] Debe tenerse en cuenta que la función de la pareja FtsZ-MreB se invirtió durante la evolución al convertirse en tubulina-actina.
Elementos del citoesqueleto de Caulobacter crescentus. En la figura, estos elementos procarióticos se relacionan con sus homólogos eucariotas y se hipotetiza su función celular.[5] Debe tenerse en cuenta que la función de la pareja FtsZ-MreB se invirtió durante la evolución al convertirse en tubulina-actina.La actinobacteria que daría lugar a los eucariontes perdió los peptidoglicanos y lipoproteínas de su pared celular y adquirió una capa S de glicoproteínas mediante la incorporación a estas moléculas de una cola hidrofóbica para su anclaje a la membrana y la N-glicosilación co-transaccional. Esta nueva superficie/pared le permitiría al organismo mantener el mecanismo de división celular bacteriana durante las primeras etapas de la transformación. En las siguientes etapas de la transformación esta nueva pared cambiaría a una superficie flexible, pero antes tuvo que desarrollarse un citoesqueleto.
La pared celular desempeña en los procariontes también un papel como cohesionador de la célula, por lo que para reemplazar esta función los eucariontes desarrollaron un primitivo citoesqueleto. Los eucariontes presentan un citosqueleto compuesto de tres proteínas fundamentales: actina, miosina y tubulina. La actina es la proteína que forma los microfilamentos y podría tener como origen el MreB de las bacterias, pues presentan similaridades en la secuencia de péptidos y en la estructura terciaria. En resumen, después de la conversión de la rígida pared celular en una superficie flexible, el MreB se convirtió en actina para actúar como un primitivo citoesqueleto. Además, la actina podría intervenir en la fagocitosis si se polimerizase para cambiar la forma de la membrana y con ello rodear parcialmente a la posible presa, incluso en ausencia de miosina.
La miosina se utiliza en los eucariontes como motor molecular para desplazar los filamentos de actina durante la división celular. En las bacterias, además de la pared celular intervienen también motores moleculares en la división celular. En concreto, las bacterias Gram-negativas utilizan MukB para la segregación del cromosoma, mientras que las bacterias Gram-positivas utilizan SMC. Estos motores moleculares serían transformados en miosina y junto con la actina darían lugar al anillo contráctil de actina-miosina de los eucariontes. Una vez que este anillo se desarrolló completamente, reemplazó al anillo de FtsZ que realiza en las bacterias esta misma función.
La tercera proteína importante del citoesqueleto de los eucariontes es la tubulina. Esta proteína tiene como origen el FtsZ bacteriano, que una vez que perdió su función en el anillo contráctil no fue abandonado del todo. El FtsZ se triplicó para formar la tubulina, centrosomas y microtúbulos del citoesqueleto de los eucariontes.
Adquisición de mitocondrias
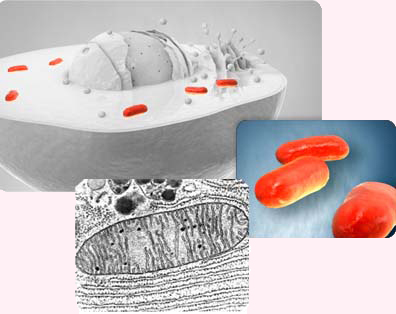
 Mitocondrias. Arriba: sección de una célula, con el núcleo en el centro y las mitocondrias coloreadas en rojo. Centro: tres mitocondrias. Abajo: Sección de una mitocondria.
Mitocondrias. Arriba: sección de una célula, con el núcleo en el centro y las mitocondrias coloreadas en rojo. Centro: tres mitocondrias. Abajo: Sección de una mitocondria.La fagocitosis también abre inmediatamente la puerta a que alguna célula ingerida escape a la digestión y se convierta en un endosimbionte. El desarrollo de la fagocitosis fue probablemente esencial para la endosimbiosis, puesto que la pared celular bacteriana actúa como una barrera para la ingestión de células. La única excepción conocida es una proteobacteria beta que contiene proteobacterias gamma en su citosol, que son transmitidas verticalmente. En cambio, una presa introducida en una vacuola digestiva podría fácilmente romper la membrana y reproducirse libremente en el citoplasma. Por tanto, fue la fagocitosis la que llevó a la endosimbiosis y no al revés.
A la aparición de la fagocitosis le debió seguir casi inmediatamente la adquisición de una proteobacteria alfa como proto-mitocondria, pues se acepta que todos los eucariontes amitocondriales anaerobios actuales descienden de antecesores aerobios con mitocondrias. Podría haber ocurrido incluso antes de que el sistema endomembranoso, citoesqueleto y núcleo estuvieran completamente formados, pero no es un paso esencial para que estos elementos fueran desarrollados. La transferencia de genes desde la proteobacteria al huésped podría haber ocurrido más fácilmente si el núcleo no se hubiese desarrollado propiamente.
A continuación se produjo la sustitución casi neutral de algunas enzimas del huésped por otras del endosimbionte, tales como la valil-ARNt sintetasa. Se conocen también otras transferencias de genes desde el segundo evento endosimbiótico primario, la adquisición de los cloroplastos, y desde los eventos endosimbióticos secundarios. La sustitución de un gen del huésped por un gen del simbionte es más fácil que en sentido contrario, pero esto último también podría haber ocurrido con algunas enzimas solubles de la mitocondria, que podrían tener un origen actinobacteriano.
Sistema de endomembranas
Véase también: Tráfico intracelular de membranasLa primitiva capacidad de los proto-eucariontes para la ingestión de presas fue poco a poco perfeccionada. Pronto se formaron endomembranas y lisosomas para constituir compartimentos estancos en los que digerir a las presas. La teoría autógena considera que el sistema de endomembranas se desarrolló a partir de invaginaciones de la membrana citoplasmática que terminaron haciéndose independientes. También se desarrolló la exocitosis para retornar las membranas a la superficie y de paso contribuir al crecimiento de la membrana citoplasmática. Algunas de las proteínas de membrana originales permanecieron embebidas en la membrana citoplasmática, mientras que otras fueron transferidas a las endomembranas. Las proteínas de membrana de las bacterias son homólogas a las de los eucariontes (tanto las de la membrana citoplasmática como las del retículo endoplasmático), por lo que se establece su origen bacteriano.
 Mediante el mecanismo de la exocitosis la célula eucariota dirige vesículas secretoras a la membrana citoplasmática. Estas vesículas contienen proteínas de membrana y lípidos que son enviadas para convertirse en componentes de la membrana, así como proteínas solubles para ser secretadas al exterior.
Mediante el mecanismo de la exocitosis la célula eucariota dirige vesículas secretoras a la membrana citoplasmática. Estas vesículas contienen proteínas de membrana y lípidos que son enviadas para convertirse en componentes de la membrana, así como proteínas solubles para ser secretadas al exterior.Paralelamente al origen de las endomembranas fue necesario el desarrollo de válvulas selectivas para diferenciar entre la membrana citoplasmática y las endomembranas. Mediante el proceso de exocitosis, las vesículas situadas en el citoplasma, cuando les llega una señal, se fusionan con la membrana, liberando su contenido. En la membrana bacteriana, la mayoría de las proteínas son insertadas o traslocadas por mecanismos post-traslacionales como el SecA, mientras que en el retículo endoplasmático eucariota la inserción es predominantemente co-traslacional. Por tanto, esto funciona como válvula selectiva que diferencia entre la membrana citoplasmática y las endomembranas y previene la secreción inútil de enzimas protolisosomales sobre la membrana citoplasmática. Con este mecanismo también se consigue que los ribosomas queden asociados permanentemente a las endomembranas y crear un primitivo retículo endoplasmático rugoso.
A continuación, las primitivas endomembranas se diferenciaron para dar lugar a los distintos compartimentos: retículo endoplasmático, aparato de Golgi, lisosomas, peroxisomas, etc. Para ello se necesitó del desarrollo de válvulas selectivas que diferenciaran entre los compartimentos. Estas válvulas se implementan mediante una citosis (gemación y fusión de vesículas) selectiva. Por ejemplo, las vesículas COP I median en el aparato de Golgi, las COP II median en el retículo endoplasmático y las clatrina lo hacen en los lisosomas. Esto previene, por ejemplo, que los receptores de los ribosomas alcancen el aparato de Golgi. Estas tres principales clases de vesículas probablemente tengan un origen común.
En resumen, a partir de las membranas de las vacuolas digestivas de la primitiva fagocitosis, se desarrolló en primer lugar la citosis de las vesículas y después se creó un sistema permanente de endomembramas.
Núcleo y mitosis
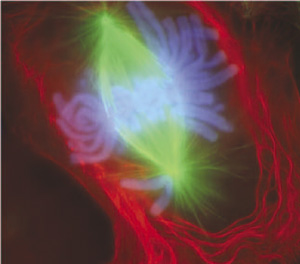
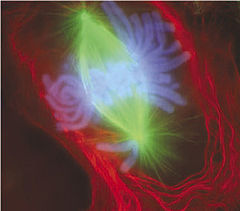 Durante la mitosis, los motores moleculares de cinesina tiran de los microtúbulos para formar el huso acromático (en verde) y así conseguir la segregación de los cromosomas (en azul).
Durante la mitosis, los motores moleculares de cinesina tiran de los microtúbulos para formar el huso acromático (en verde) y así conseguir la segregación de los cromosomas (en azul).La membrana nuclear es bastante más efectiva para la protección del ADN que la superficie de cromatina de las bacterias. Probablemente se desarrolló para evitar los daños que podrían causar en el ADN los nuevos motores moleculares de miosina, dineína y cinesina durante la fagocitosis, citocinesis y el transporte de vesículas. Por tanto, los cambios en la organización del genoma serían a su vez causados por los otros cambios en cascada que tuvieron lugar por la adquisición de la fagocitosis.
El genoma queda protegido en los cromosomas por el enrollado del ADN durante la interfase e incluso durante la mitosis el enrollado es todavía más compacto. Además, se añadieron las nuevas histonas H2a y H2b a las histonas H3 y H4 desarrolladas en el antecesor neomura y a la H1 presente ya en el antecesor actinobacteriano. También se desarrolló la acetilación reversible de las histonas para mediar en la compactación mitótica, probablemente antes que la membrana nuclear.
El origen del núcleo está íntimamente ligado a la mitosis, teniendo en cuenta que los cromosomas están rodeados de endomembranas que dificultan su segregación. La mitosis de los eucariontes es un proceso traumático en donde los microtúbulos tiran fuertemente de los cromosomas. Tampoco la segregación del cromosoma en las bacterias es un proceso puramente pasivo sino que también envuelve motores moleculares. Durante la segregación del ADN, las bacterias Gram-positivas, al igual que los eucariontes, usan la familia de proteínas MukB, mientras que las bacterias Gram-negativas usan la familia SMC. Los motores de proteínas bacterianos MukB/SMC pueden haber sido los antecesores de la miosina, dineína y cinesina de los eucariontes.
Además, durante la división celular de las bacterias, la proteína FtsZ se ensambla en un anillo contráctil para mediar en la división, al igual que lo hace en la división de los cloroplastos y mitocondrias más primitivas. FtsZ dejó esta función de anillo contráctil cuando se transformó en la tubulina de los eucariontes. Un nuevo anillo contráctil de actina-miosina se desarrolló en los eucariontes para realizar la antigua función del FTsZ.
Complejo de poros nucleares
 Poro nuclear. Vista lateral 1. Envoltura nuclear. 2. Anillo externo 3. Rayos. 4. Canasto. 5. Filamentos. (Dibujado en base a microscopía electrónica).
Poro nuclear. Vista lateral 1. Envoltura nuclear. 2. Anillo externo 3. Rayos. 4. Canasto. 5. Filamentos. (Dibujado en base a microscopía electrónica).La función primaria del complejo de poros nucleares, grandes complejos de proteínas que atraviesan la membrana nuclear, probablemente fuera evitar la fusión total de vesículas en una doble membrana continua que aislara el núcleo del citoplasma. Inicialmente tendrían poros muy grandes y serían una ruta pasiva que permitía el movimiento de moléculas solubles entre el citoplasma y el nucleoplasma. Antes que la ruta pasiva se cerrara totalmente, el mecanismo activo de entrada/salida tuvo que desarrollarse con suficiente eficacia. Probablemente el objetivo inicial del mecanismo activo hubiese sido simplemente acelerar el intercambio y con ello aumentar la velocidad de crecimiento de la célula.
Reproducción sexual
La pérdida de la pared celular podría haber permitido la reproducción sexual muy tempranamente y que no haya habido eucariontes primitivamente asexuales. El citoesqueleto, los motores moleculares y el sistema de endomembranas también facilitan la reproducción sexual.
La fusión de células esta relativamente extendida entre los protistas. La mayoría tienen también fusión nuclear y meiosis, mientras que otros, denominados agámicos, no las tienen, por ejemplo, el alga cercozoa Chlorarachnion y el haptophyta Reticulosphaera. La fusión de células podría haberse desarrollado sin que el objetivo fuese reproducción sexual. Por ejemplo, los plasmodios, grandes células multinucledas que se producen entre los hongos mucilaginosos, tienen como objetivo la búsqueda del alimento.
Una fusión nuclear accidental o un fallo en la mitosis harían un núcleo poliploide. La meiosis podría haberse desarrollado para reparar estos errores. Esto es, podría haber habido una fase intermedia con fusión de células y meiosis pero sin fusión nuclear.
Referencias
- ↑ a b T. Cavalier-Smith (2002). «The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa». International Journal of Systematic and Evolutionary Microbiology 52: pp. 297-354.
- ↑ Thomas Cavalier-Smith (2002), The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification, International Journal of Systematic and Evolutionary Microbiology 52: 7–76
- ↑ Cavalier-Smith T (2006). «Cell evolution and Earth history: stasis and revolution». Philos Trans R Soc Lond B Biol Sci 361 (1470): pp. 969–1006. PMID 16754610. http://www.journals.royalsoc.ac.uk/content/0164755512w92302/fulltext.pdf.
- ↑ Thomas Cavalier-Smith (2006), Rooting the tree of life by transition analyses, Biol Direct. 1: 19. doi: 10.1186/1745-6150-1-19.
- ↑ Gitai, Z. (2005). «The New Bacterial Cell Biology: Moving Parts and Subcellular Architecture». Cell 120 (5): pp. 577-586. doi:. http://linkinghub.elsevier.com/retrieve/pii/S0092867405001935.
Véase también
- Teoría autógena
- Teoría endosimbiótica


Wikimedia foundation. 2010.