- Quimiotaxis
-
Quimiotaxis
La quimiotaxis es un tipo de taxis, un fenómeno en el cual las bacterias y otras células de organismos uni o multicelulares dirigen sus movimientos de acuerdo a ciertas sustancias químicas en su medio ambiente.
Esto es importante para que las bacterias encuentren alimento (por ejemplo, glucosa) nadando hacia la mayor concentración de moléculas alimentarías o para escapar de venenos (p. ej.: fenol).
En los organismos multicelulares la quimotaxis es fundamental tanto en las fases tempranas del desarrollo (como los movimientos de espermatozoides hacia el óvulo durante la fertilización) como en las fases más tardías (como la migración de neuronas o linfocitos) como para las funciones normales.
Además se ha reconocido que los mecanismos que permiten la quimiotaxis en animales pueden ser destruidos durante las metástasis del cáncer.
La quimiotaxis se denomina positiva si el movimiento es en dirección hacia la mayor concentración de la sustancia química en cuestión y negativa si es en dirección opuesta.
Contenido
Historia de la investigación de la quimiotaxis
Aúnque la migración celular fue detectada desde los primeros días del desarrollo del microscopio (Lee Wenhack) la primera descripción erudita de quimiotaxis fue hecha por primera vez por T.W. Ergelmann (1881) y W.F. Pjeffer. (1884) en bacterias y H.S. Jennings (1906) en ciliados.
El Premio Nobel Laurate E. Mechnikov, también contribuyó al estudio con sus investigaciones de los procesos en los pasos iniciales de la fagocitosis.
La importancia de la quimiotaxis en la biología y en la patología clínica fue ampliamente aceptada en la década de 1930. Las definiciones más fundamentales asociadas a los fenómenos fueron también redactadas en ese tiempo.
Los aspectos más importantes en el control de calidad de las técnicas para estudiar la quimiotaxis (chemotaxis assays) fueron descritos por H. Harris en los años 1950.
Entre 1960-1970, la revolución de la biología celular y bioquímica moderna descubrió una serie de nuevas técnicas que hicieron posible investigar las células migratorias y las fracciones subcelulares responsables del movimiento de estas.
El trabajo pionero de J. Adler, representa un punto de cambio significativo en la comprensión de todo el proceso de la traducción de señales intracelulares de las bacterias.[1]
El 3 de Noviembre de 2006, el Dr. Dennis Breins de la Universidad de Cambridge fue premiado con la condecoración del Premio de Ciencias de Microsoft Europea por su trabajo en Quimiotaxis E. Coli.[2] [3]
Filogenia y señalización quimiotáctica
La quimiotaxis es una de las respuestas fisiológicas más básicas de la célula.
El desarrollo del sistema de receptores para la detección de sustancias, dañinas y favorables, en el medio ambiente fue lo más esencial para los organismos unicelulares desde las etapas más tempranas de la filogenia.
El análisis comprensivo de la actividad quimiotáctica del protozoo Tetrahymena pyriformis y las consecuentes secuencias en la aparición de aminoácidos en el caldo primario, sugiere que habría una buena correlación entre el carácter quimiotáctico de estas relativamente simples moléculas orgánicas y su desarrollo en la tierra.
De esta manera las primeras moléculas sugieren ser altamente quimioatrayentes (p. ejemplo Gly, Glu, Pro), mientras que tarde se encontraron algunos pensaron que son fuertemente quimiorepelentes (p.ej.: Tyr, Trp, Phe ) a los aminoácidos.[4]
Quimiotaxis bacteriana
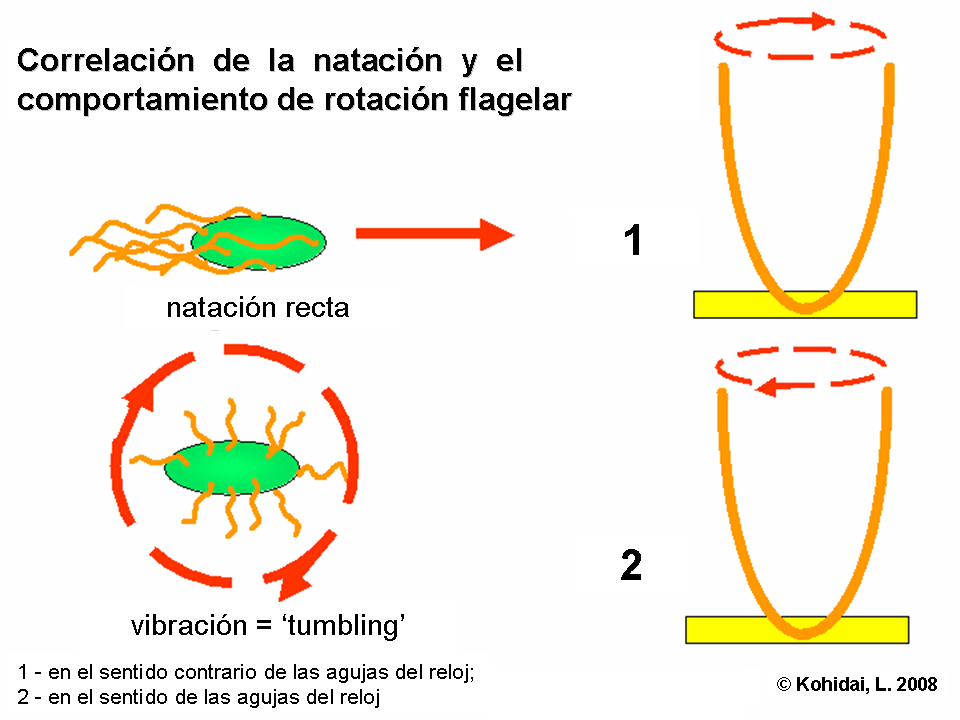
Algunas bacterias como la E. coli poseen muchos flagelos por célula (habitualmente entre 4 y 10). Estas pueden rotar en dos sentidos:
- Rotación en el sentido contrario a las agujas del reloj, lo cual alinea los flagelos en un haz de rotación simple, causando que la bacteria nade en línea recta.
- Rotación en el sentido de las agujas del reloj, dobla el flagelos en un haz haciendo que cada flagelo apunte en dirección diferente causando detención del movimiento y dejando la bacteria en un lugar.
La dirección de rotación es dada por un observador externo, mirando desde abajo hacia la cédula.
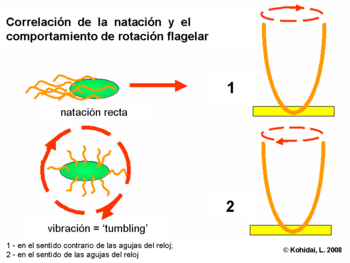
Conducta
El movimiento total de la bacteria es el resultado de la alternancia entre las fases de detención y natatorias. Si alguien observa la bacteria nadando en un entorno uniforme, el movimiento se ve como un paseo arbitrario, con un nado en línea recta interrumpido con detenciones arbitrarias que reorientan a la bacteria. Las bacterias como E. coli son incapaces de elegir la dirección en la cual nadan, y son incapaces de nadar en una línea recta por más de algunos segundos, debido a la difusión rotatoria. En otras palabras, las bacterias olvidan la dirección a la cual se dirigen. Dadas estas limitaciones, es extraordinario que las bacterias puedan dirigir sus movimientos y encontrar lugares favorables de alta concentración de atrayentes (habitualmente alimentos) y evadir los repelentes (habitualmente venenos).
En la presencia de gradientes químicos, la bacteria realiza quimiotaxis o dirige su movilidad arbitraria basada en el gradiente. Si la bacteria siente que su movimiento va en la dirección correcta (hacia el atrayente y lejos del repelente) mantendrá su natación en una línea recta por un tiempo más largo antes de detenerse. Si su movimiento va en una mala dirección, se detendrá más rápido e intentará una nueva dirección al azar (aleatoria). En otras palabras, una bacteria como E. coli usa sus sensibilidad temporal para decidir si la vida esta mejorando o empeorando. De esta manera si encuentra la ubicación con mayor concentración de atrayentes (usualmente la fuente) es mejor. Incluso a altas concentraciones es capaz de distinguir hasta las más pequeñas diferencias en concentraciones (atrayentes/repelentes). La función de escapar de los repelentes funciona con la misma eficiencia.
Parece considerable que este movimiento uniforme con propósito, es el resultado de una simple elección entre dos métodos de movimientos aleatorios, llamados detención y natación línea recta. De hecho, las respuestas quimiotácticas como el olvido de la dirección y la selección del movimiento, se parecen a las habilidades de tomar decisiones, en las superiores formas superiores de vida, que procesan los datos sensoriales con cerebros.
La naturaleza helical del filamento flagelar individual es critico, para que este movimiento ocurra. Así, como la proteína que forma el filamento flagelar, el flagelin es similar entre todas las bacterias flageladas. Los vertebrados parecen tener ventaja en este hecho al poseer un inmunoreceptor (TLR5) desigñado para reconocer la proteína conservada.
Así como hay muchas instancias en la biología, hay bacterias que no siguen estas reglas. Muchas bacterias, como es el Vibrio, son monoflageladas y tienen este único flagelo en un polo de la célula. Su método de quimiotaxis es diferente. Otros poseen un solo flagelo que se mantiene dentro de la pared de la célula. Esas bacterias se mueven a través de un rodamiento de toda la célula, lo cual se parece al movimiento del sacacorchos.[5]
Transducción de la señal
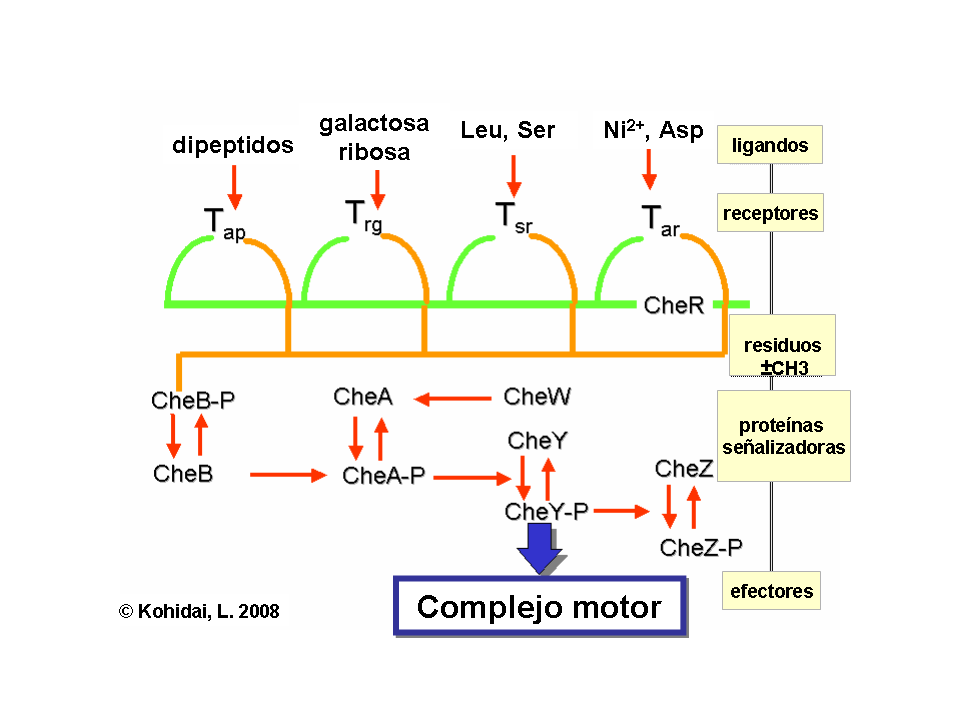
Los gradientes químicos son detectados a través de muchos receptores transmembranales llamados "methyl accepting chemotaxis proteins" (proteínas quemotácticas que aceptan grupos metilo) (MCPs) las cuales varían en las moléculas que ellas detectan. Estos receptores pueden unir atrayentes o repelentes directa o indirectamente a través de la interacción con proteínas del espacio periplasmatico. Las señales de estos receptores son transmitidas a través de la membrana plasmática hacia el citosol, donde las Che proteínas son activadas. Las Che proteínas cambian las frecuencias de avance y giro, y alteran los receptores.
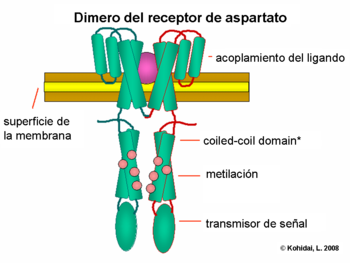
Regulación de los flagelos
Las proteínas CheW y CheA se unen al receptor. La ausencia de atrayentes causa una auto fosforilación en la histidinaquinasa, CheA, a través del único residuo de histidina altamente conservado. CheA, a la vez transfiere grupos forforilados para conservar residuos de aspartato en respuesta de los reguladores CheB y CheY [nota: CheA es una histidinaquinasa y no transfiere activamente grupos forforilados. La respuesta reguladora de CheB, toma el grupo fosforilado de CheA]. Este mecanismo de traducción de señal se llama “Two Component System” (Sistema de dos componentes) y es la forma común de la traducción de señal en las bacterias. CheY induce la detención del movimiento a través de la interacción con la proteína flagelar-interruptora proteína FliM, induciendo un cambio en el sentido de la rotación del flagelo, desde el sentido contrario a las agujas del reloj hacia el sentido de las agujas del reloj. El cambio del estado de la rotación de un solo flagelo es capaz de interrumpir el haz completo bulto y causar una vibración.
Regulación de receptores
CheB, una vez activado por CheA, actúa como una metilesterasa, removiendo grupos metilo desde los residuos de glutamato en el lado citosólico (intracelular) del receptor. Esto trabaja en forma antagonista con la CheR, una metiltransferasa que adjunta residuos de metilo a los mismos residuos de glutamato. Mientras más residuos metilo se unen al receptor, mas aumenta la sensibilidad del receptor. Una vez se detecta presencia de atrayente, se inhibe la autofosforilación de CheA, y por tanto de CheB, por lo que se induce una desmetilación del receptor. Del mismo modo, la regulación por retroalimentación (feedback) ajusta la metilación continuamente a los niveles ambientales, manteniendo sensibilidad para los más leves cambios del medio ambiente, incluso con concentraciones químicas restantes extremadamente bajas. Esta regulación permite a la bacteria, recordar las concentraciones químicas desde el pasado reciente y compararlas con aquellas que corrientemente experimenta (es decir, compara la presencia de señales extracelulares con la presencia de las mismas momentos antes, la cual es recordada por el nivel de metilación). Este “conocimiento” le da la posibilidad de hacer el viaje contra o a favor del gradiente. No obstante, el sistema de metilación sólo no puede explicar la amplia gama de sensibilidad que las bacterias poseen hacia los gradientes químicos. Mecanismos reguladores adicionales, como la agrupación de receptores y la interacción receptor-receptor también modula la vía de señalización
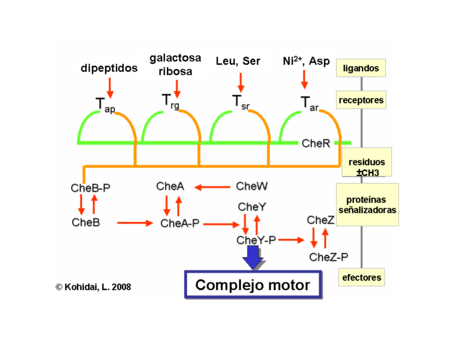
Quimiotaxis eucariota
El mecanismo, a través del cual, las células eucariotas realizan la quimiotaxis es diferente a las de las bacterias. Sin embargo, la sensibilidad de los gradientes químicos sigue siendo un paso crucial en el proceso. Debido a su tamaño, las procariotas no pueden detectar en forma efectiva la concentración de gradientes, por lo cual, estas células buscan y evalúan su entorno a través de una natación constante (pasos consecutivos de natación recta y detenciones, sin un movimiento desplazado). Al contrario de las procariotas, el tamaño de las células eucariotas permiten la posibilidad de detectar el gradiente lo cual resulta en una distribución de receptores dinámica y polarizada. La inducción de esos receptores a través de quimioatrayentes y quimiorepelentes resulta en una migración para alejarse o acercarse a las sustancias quimiotacticas.
Los niveles de receptores, las vías de las señalización intracelular y los mecanismos efectores, todos representan componentes típicos de los eucariotas. En las células eucariotas unicelulares los movimientos ameboides y los cilios o los flagelos eucarióticos son los principales efectores (p.ej.: Amoeba o Tetrahymena). Algunas células eucariotas cuyo es de vertebrados superiores, así como células inmunes, también se mueven a donde ellas necesitan. Detrás de las células inmunes competentes como (granulocitos, monolitos y linfocitos) se encuentran un grupo grande de células - consideradas propias y fijas de los tejidos - que también son móviles en condiciones especiales, sean estas fisiológicas (p.ej.: mastocitos, fibroblastos, células endoteliales) o patológicas (p.ej.: metástasis )respectivamente.
La quimiotaxis tiene un significado tanto en las fases tempranas de la embriogenesis como en el desarrollo de capas germinales que es dirigido por los gradientes de moléculas de señal.
Motilidad
A diferencia de la movilidad en la quimiotaxis bacteriana, el mecanismo del movimiento físico a través del cual se moviliza la célula eucariota, no está claro. Aparentemente los mecanismos por los cuales los gradientes externos quimiotácticos son detectados y convertidos al gradiente PIP3 intracelular, cuyo resultado del gradiente es la activación de la vía de señalización, culminaría en una polimerización de las actinas de filamentos. El crecimiento distal del final del filamento de actina desarrolla conexiones con la superficie interna de la membrana plasmática, a través de diferentes tipos de péptidos dando resultado a una formación de pseudópodos. Los cilios de las células eucariotas también pueden dar como resultado la quimiotaxis, aún cuano en este caso es principalmente una inducción de el sistema microtubular del cuerpo basal dependiente de Ca2+ y del rompimiento microtubular de los cilios 9x2+2.
El golpe orquestado de cientos de cilias es sincronizado por un sistema submembranoso construido entre los cuerpos básales. Los detalles de la vía de señalización aún no están totalmente aclarados.
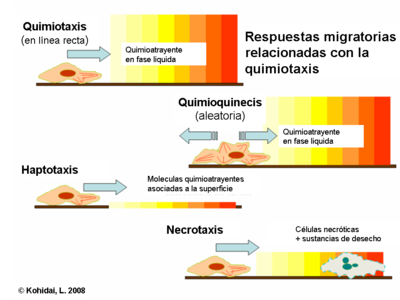
Respuestas migratorias relacionadas con quimiotaxis
Aunque la quimiotaxis es la forma de migración más frecuentemente estudiada hay muchas otras formas de movilidad a nivel celular.
- Quimioquinesis: también inducida por moléculas de la fase liquida del medio ambiente rodeante. Sin embargo la respuesta obtenida no es vectorial. Ni la frecuencia ni la amplitud de este movimiento tiene carácter direccional, los componentes de este comportamiento más bien sirven para percibir el medioambiente (palpándolo), más que la búsqueda de la migración entre dos diferentes puntos.
- Haptotaxis: el gradiente de los quimioatrayentes es expresado sobre una superficie o unido a esta, en contraste a la vía clásica de la quimiotaxis cuando el gradiente se desarrolla en un espacio soluble. La principal superficie biológicamente activa del mecanismo haptotaxico es la matriz extracelular (ECM), la presencia de ligandos unidos es responsable de la inducción de la migración y angiogénesis transendotelial.
- Necrotaxis: encarna un tipo especial de quimiotaxis en el cual la molécula quimioatrayente esta liberada de la célula necrótica o apoptótica (muerta). Dependiendo del carácter químico de las sustancias liberadas, la necrotaxis puede acumular o rechazar células, lo que subraya el significado fisiopatológico de este fenómeno.
Receptores
La mayoría de las células eucariotas detectan los estímulos quimiotácticos es a través de receptores con siete dominios transmembranales acoplados a proteínas G heterotriméricas. Esta clase de receptores es grande, representan una parte significativa del genoma. Algunos miembros de esta super familia de genes, son usados en la visión tanto como en el olfato (rodopsina y odoríferos respectivamente). Las principales clases de receptores quimiotácticos profesionales se disparan por los formil peptidos, los llamados "formil peptid receptors" (FPR), las quimioquinas o receptores de quimioquinas (CCR o CXCR) y leucotrienos, receptores de leucotrienos (BLT) Sin embargo, la inducción de un amplio conjunto de receptores de membrana (p.ej.: aminoácidos, insulina, peptidos vasoactivos), también producen la migración de la célula.
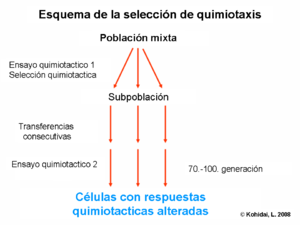
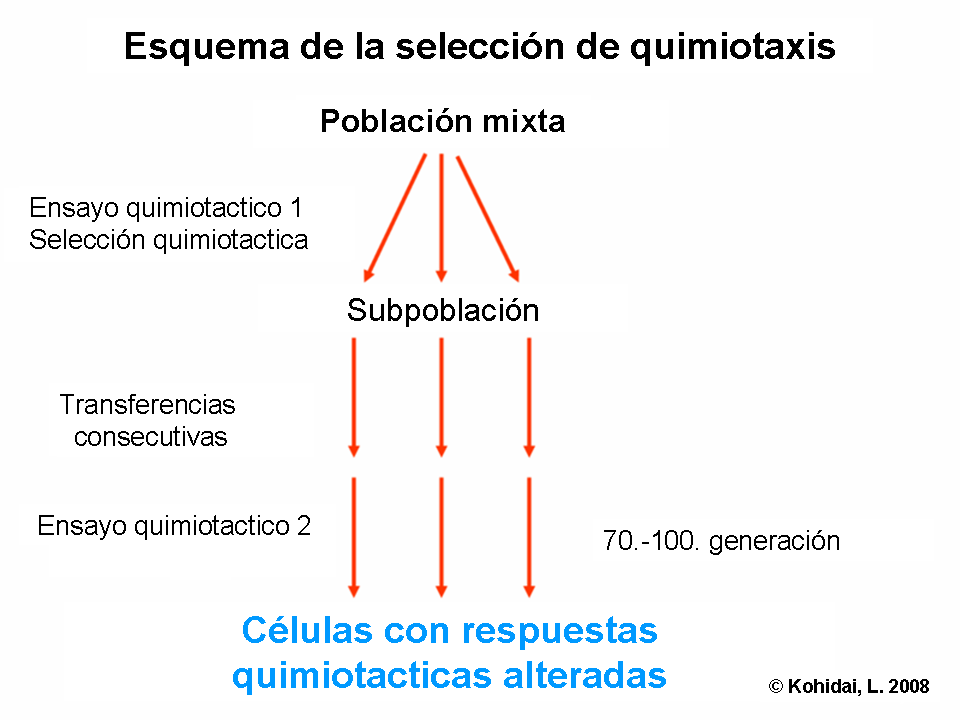
Seleccción quimiotáctica
Aunque algunos receptores quimiotacticos se expresan en la superficie de la membrana con características de largo plazo que están genéticamente determinadas, otros tienen una dinámica a corto plazo, ya que se unen al ligando, que permite la posibilidad de selección de células receptoras quimiotacticas con un ensayo simple de quimiotaxis. A través de la selección quimiotáctica podemos determinar también, si una molécula indeterminada (no caracterizada) actúa vía camino de larga o corta duración. El término selección quimiotáctica es también usado para designar la técnica la cual separa a la células eucariotas o procariotas de acuerdo a sus ligandos electores quimiotácticos responsables del movimiento.[6]
Ligandos quimiotácticos
El número de moléculas capaces de obtener una respuesta quimiotáctica es relativamente alta y nosotros somos capaces de distinguir moléculas quimiotácticas primarias y secundarias:
El principal grupo de ligandos primarios son los siguientes:
- Formil péptidos son, di-,tri- tetrapeptidos de origen bacteriano (véase grupo formil en la porción amino terminal del peptido) estas son obtenidas desde las bacterias en vivo o después de la descomposición de la célula. Un típico miembro de este grupo es el N-formilmetionil-leucil-fenilalanina (fMLF or fMLP en las referencias). Las de origen bacteriano fMLF son componentes claves en la inflamación y tienen características de efectos quimioatrayentes en los granulocitos neutrofilos y monocitos.
- Complemento 3 a (C3a) y complemento 5a (C5a) son productos intermedios de la cascada del complemento. Su síntesis esta relacionada con tres vias (clásica, dependiente de lectinas y la alternativa) de activación del complemento a través de la enzima convertasa. La principal célula blanco de estos derivados, son tanto los granulocitos neutrofilos como los monocitos.
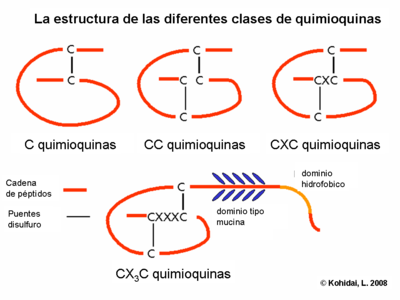
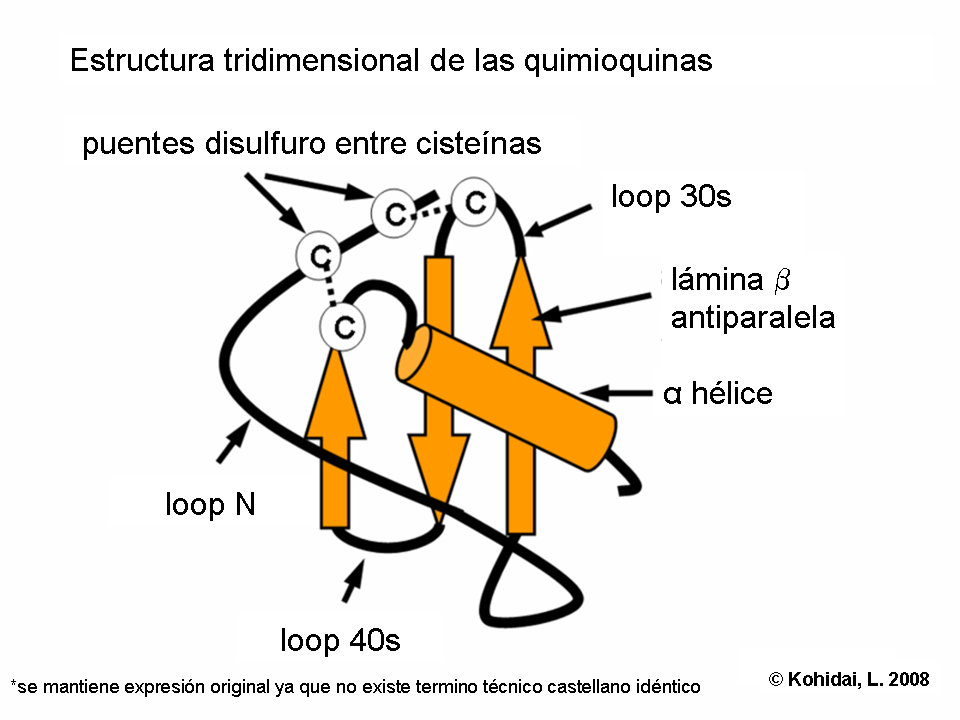
- Quimiocinas, pertenecen a una clase especial de citoquinas. Sus grupos (C, CC, CXC, CX3C, quimioquinas) representan no solamente moléculas estructuralmente relacionadas con un especial arreglo de puentes disulfuro, sino también la especificidad de sus células objetivos. Son diversos: las CC quimioquinas actúan en monocitos (ejemplo en RANTES), CXC quimioquinas son específicas de granulocitos (neutrofilos) (ejemlo IL-8).
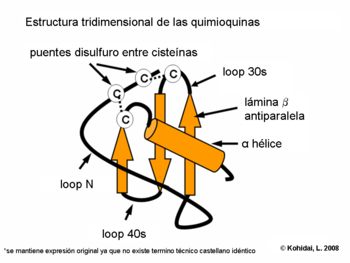
Las investigaciones de la estructura tridimensional de las quimoquinas prueban que la composición de hoja beta y alpha hélice proporciona la expresión de las secuencias necesarias para la interacción con los receptores de quimioquinas. La formación de dímeros y su actividad biológica aumentada fue demostrado por cristalografía de varias quimioquinas (p.ej.: IL-8)
- Los leucotrienos pertenecen al grupo de eicosanoides. Son mediadores lipídicos importantes de la cascada del ácido araquidónico convertidos por la 5-lipoxigenas. Su miembro predominante es el leucotrieno B4 (LTB4) los cuales obtienen adhesión, quimiotaxis y agregación de los leucocitos. El efecto quimioatrayente del LTB4 se caracteriza por ser inducido por un receptor de siete dominios transmembranales y que esta acoplado a una proteína G. Estos son altamente son altamente expresados en la inflamación y la alergia.
Rangos quimiotacticos efectivos (quimiotactic range fitting CRF)
La respuesta quimiotactica generada por la interacción ligando receptor se distingue generalmente por unas concentraciones óptimas efectivas de ligandos. Sin embargo la correlación de la amplitud y radio provocada por las células que responden comparadas con el número total, es también característico de las señales quimiotacticas. Las investigaciones de las familias de ligandos (ejemplo aminoácidos y oligopeptidos) prueban que los rangos efectivos (amplitudes; números de células objetos) y las actividades quimiotacticas dependen de rangos efectivos: la fracción de quimioatrayentes acompañado por rangos amplios, mientras que el carácter quimiorepelente se caracteriza por rangos estrechos de ligandos.
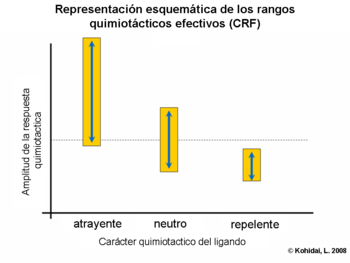
Importancia clínica
El cambio del potencial migratorio de las células tiene una importancia relativamente alta en el desarrollo de varios síntomas y síndromes clínicos. La alteración de la actividad quimiotáctica extracelular o intracelular de patógenos, como por ejemplo: Escherichia coli; Listeria monocytogenes respectivamente, en si mismo representa un objetivo clínico significativo. La modificación de la habilidad quimiotactica endógena de estos microorganismos por agentes farmacéuticos pueden disminuir o inhibir el radio de acción de las infecciones o la difusión de estas enfermedades infecciosas. Aparte de estas infecciones, hay otras enfermedades donde la quimiotaxis alterada es el factor etiológico primario, como es el síndrome Chediak-Higashi donde las gigantes vesículas intracelulares inhiben la normal migración de las células.
Quimiotaxis en las enfermedades Tipo de enfermedad Quimiotaxis aumentada Quimiotaxis disminuida Infecciones Inflamaciones SIDA, brucellosis La quimiotaxis es el resultado de la enfermedad - Síndrome Chediak-Higashi, Síndrome Kartagener Quimiotaxi es afectada Artereosclerosis, artritis, periodontitis, psoriasis, injuria de reperjusion, tumores metastasicos Esclerosis múltiples, Enfermedad de Hodgkin, Infertilidad masculina Intoxicaciones asbestos, benzopireno Sales de mercurio y cromo, ozono (O3) Medición de la quimiotaxis (cuantificacion de la quimiotaxis)
Un alto rango de técnicas están disponibles para la evaluación de la actividad quimiotaxica de la célula o del carácter quimioatrayente o quimiorepelente de los ligandos. Los requerimientos básicos de la cuantificación son los siguientes:
- La concentración de gradientes pueda desarrollarse relativamente rápido y persistir por mucho tiempo en el sistema.
- Se distinguen las actividades quimiotacticas y quimiocineticas
- La migración de células es libre hacia fuera y sobre el eje del la gradiente de concentración.
- Las respuestas detectadas son el resultado de la migración activa de células.
A pesar que un ensayo ideal de medición de quimiotaxis no esta disponible, existen varios protocolos y equipos los cuales pueden ofrecer una buena oferta con las condiciones que se describen más abajo. Las más comúnmente usadas son:
- Ensayos de placa de agar (ejemplo PP-chamber)
- Técnicas de dos cámaras (Two chamber techniques) (ejemplo Boyden-Chamber-Zigmond Chamber-Dunn Chamber-Multi-Well Chambers- Capillary Techniques)
- Otros (ejemplo T-maze technique-Opalescence technique-Orientation assays)
Un capítulo más detallado se puede encontrar bajo el título de ensayos de quimiotaxis.
Referencias
- ↑ Julius Adler and Wung-Wai Tso (1974). «Decision-Making in Bacteria: Chemotactic Response of Escherichia Coli to Conflicting Stimuli» Science. Vol. 184. pp. 1292–4.
- ↑ http://research.microsoft.com/displayArticle.aspx?id=1572 retrieved November 6, 2006
- ↑ http://news.bbc.co.uk/2/hi/science/nature/6113522.stm retrieved November 6, 2006
- ↑ The 'Optimal' Chemotactic Ligand - Amino acids retrieved February 3, 2008
- ↑ Howard C. Berg (2003). «E. coli in motion» Springer-Verlag, NY.. Vol. ISBN 0-387-00888-8.
- ↑ Kohidai L and Csaba G (1988). «Chemotaxis and chemotactic selection induced with cytokines (IL-8, RANTES and TNF alpha) in the unicellular Tetrahymena pyriformis.» Cytokine. Vol. 10. pp. 481–6.
Enlaces externos
 Wikilibros alberga un libro o manual sobre Quimiotaxis.
Wikilibros alberga un libro o manual sobre Quimiotaxis.
 Wikimedia Commons alberga contenido multimedia sobre Quimiotaxis.
Wikimedia Commons alberga contenido multimedia sobre Quimiotaxis.
- Chemotaxis
- Cell Migration Gateway
- Global Existence for Chemotaxis with Finite Sampling Radius
- Bacterial Chemotaxis Depends on a Two-Component Signaling Pathway Activated by Histidine-Kinase-associated Receptors, Molecular Biology of the Cell 4th Edition © 2002 by Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter.
- Figure 15-69. The two-component signaling pathway that enables chemotaxis receptors to control the flagellar motor during bacterial chemotaxis, Molecular Biology of the Cell 4th Edition © 2002 by Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter.
Categorías: Motilidad celular | Sistema inmunológico




Wikimedia foundation. 2010.