- Sincitio
-
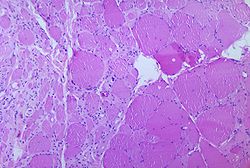 Micrografía de músculo esquelético. En la periferia de cada célula cortada transversalmente (con forma aproximadamente circular) pueden verse varios núcleos.
Micrografía de músculo esquelético. En la periferia de cada célula cortada transversalmente (con forma aproximadamente circular) pueden verse varios núcleos.
En biología, un sincitio,[1] sincicio[2] o cenocito[3] [4] es una célula que contiene muchos núcleos.[5] La mayoría de las células en los organismos eucariotas de plantas y animales tienen un solo núcleo;[6] los sincitios son formas especiales. Sin embargo, en algunos organismos no solo son estructuras habituales sino que, incluso, constituyen el estado más preponderante del ciclo vital —ciertos nematodos o mixomicetos, entre otros—.
El sincitio puede formarse tanto por fusión de células individuales —en el caso del músculo estriado en los mamíferos— como por división celular incompleta —en insectos, por ejemplo—.[7] La función del sincitio es variada y depende del organismo evaluado, aunque en general puede afirmarse que funciona como una sola unidad coordinada compuesta de varias celdas vinculadas estructural y funcionalmente, que permiten el intercambio de moléculas e impulsos eléctricos. Los sincitios, además de ser el resultado del plan de desarrollo normal de un organismo, pueden ser la consecuencia de procesos infecciosos ocasionados por virus y otros patógenos.
Contenido
Formación
En organismos unicelulares los sincitios pueden formarse de dos formas. La primera (menos frecuente) es la fusión del protoplasma (pero no de los núcleos) de células ameboideas independientes entre sí pero de una misma especie. El resultado de esta fusión es una formación que recibe el nombre de plasmodio. Un ejemplo lo representan los hongos mixomicetos. La forma más común de formación de sincitios es el crecimiento de la célula primitiva a medida que su núcleo se divide, pero sin segmentar el protoplasma. El resultado es un organismo que puede ser considerado tanto como una única célula polinucleada o un organismo pluricelular. Algunas especies de protozoos (como el infusorio Opalina ranarum y numerosas especies de radiolarios) y talofitas (entre los que se incluyen las algas sifonales y los mohos mucilaginosos) son las únicas que poseen organismos puramente sinciciales.[8]
En los organismos pluricelulares, un sincitio puede formarse por fusión o por división celular. Muchos insectos, como Drosophila melanogaster, ponen huevos que inicialmente se desarrollan como blastodermos sincitiales, es decir, a principios de la formación del embrión muestran una división celular incompleta, separando los núcleos y volviendo a ensamblarse pero sin citocinesis. Por lo tanto, los núcleos se multiplican en un espacio citoplasmático común.[7] En cambio, las grandes fibras del músculo esquelético se forman por fusión de células musculares individuales.[9] Los mioblastos (células originadoras de las fibras musculares)[10] sufren un proceso de diferenciación celular dirigido por factores de transcripción como MyoD[11] que los obliga a proliferar en presencia de factores de crecimiento. Cuando ya no hay IGF, los mioblastos dejan de proliferar y comienzan a fusionarse, formando una sola célula con múltiples núcleos.[7]
Algas
 Caulerpa prolifera, un alga verde cuyo talo está constituido por un sincitio, representa las células más grandes que se conocen.
Caulerpa prolifera, un alga verde cuyo talo está constituido por un sincitio, representa las células más grandes que se conocen.Entre las algas hay varias especies cuyos organismos son enteramente cenocitos, es decir, que están formados por una sola célula gigantesca y multinucleada. Caulerpa, a modo de ejemplo, es un género de algas verdes perteneciente a la familia Caulerpaceae, cuyos integrantes consisten en una sola célula que puede llegar a medir hasta 3 metros, lo que las convierte en las células vivientes más grandes del mundo.[12] [13]
Las células de las algas del género Chlorophyta son sincitios contenidos por la pared celular que se convierte en células uninucleares al dividirse.[14] En forma inversa, las algas sifonocladales o cladoforales como Acetabularia comienzan su vida en forma unicelular pero luego se convierten en cenocíticas.[15] El talo de Siphonocladus pusillus, por ejemplo, tiene un aspecto similar a una vesícula cuya pared celular está revestida por una capa multinucleada de protoplasma y en el centro se ubica una vacuola. A medida que el ejemplar envejece, el citoplasma comienza a desprenderse formando pequeñas vesículas con vacuolas recubiertas por una capa de protoplasma polinuclear que se fragmentó de la capa original. Estas pequeñas vacuolas desarrollan una pared de celulosa mientras flotan en el interior de la vacuola central, cuya apariencia se vuelve similar a la de un filamento con tabiques que crecen ramificándose lateralmente, fenómeno conocido como división segregativa y que es característico de este tipo de algas. En cladoforáceas como Chaetomorpha o Cladophora el proceso es algo distinto. En forma independiente de la división nuclear, el protoplasma se divide en dos y se separa de la pared celular, formándose en esa zona un anillo de membrana hacia el centro que crea un tabique transversal doble que se une por cada lado con la capa interna de la pared longitudinal.[16]
Mohos mucilaginosos
 Durante una parte de su ciclo vital llamada plasmodio, los mixomicetos forman sincitios. El plasmodio de Physarium polycephalum por ejemplo, constituye una única célula con millones de núcleos que tienen la capacidad de dividirse en forma sincrónica para convertirse en esporas con un solo núcleo cada una.
Durante una parte de su ciclo vital llamada plasmodio, los mixomicetos forman sincitios. El plasmodio de Physarium polycephalum por ejemplo, constituye una única célula con millones de núcleos que tienen la capacidad de dividirse en forma sincrónica para convertirse en esporas con un solo núcleo cada una.Los mixomicetos son un grupo peculiar de mohos mucilaginosos de distribución cosmopolita que pueden crecer en el suelo húmedo y oscuro de los bosques o en la madera que se descompone luego de humedecerse por las lluvias. Durante su ciclo de vida toman tres formas distintas: mixoameba, moho mucilaginoso y espora. En su etapa primitiva o de mixoamebas, son amebas uninucleadas, haploides, sin pared celular, que se reproducen asexualmente mediante mitosis y se mueven mediante seudópodos o flagelos. Las mixoamebas pueden seguir proliferando de forma indefinida mientras haya nutrientes y el medio ambiente continúe siendo favorable. Cuando las condiciones se vuelven adversas, forman masas gelatinosas que se deslizan lentamente por el suelo. En esta etapa normalmente se denominan mohos mucilaginosos. El modo en que este moho se desarrolla a partir de las mixoamebas varía dependiendo del grupo de mixomiceto considerado. En el grupo de los mixomicetos plasmodiales (o acelulares) del orden Myxogastria, las mixoamebas actúan como gametos fusionándose entre sí para originar un cigoto, cuyo núcleo se divide repetidamente de forma tal que la célula se constituye en un sincitio, es decir, en una gran masa protoplasmática y multinucleada sin ningún tipo de pared, denominada plasmodio. Por lo general el tamaño de estos plasmodios, que pueden contener millares de núcleos, es de algunos centímetros, aunque en algunos casos pueden alcanzar un área de hasta 2 m2. Muchos tienen colores brillantes tales como amarillo, marrón o blanco. Bajo condiciones de sequedad, se recogen en una masa de la que surgen los cuerpos fructíferos que forma esporas, superficialmente similares a los esporocarpos de los hongos. Las esporas originan gametos biflagelados o ameboides que se funden en parejas para producir nuevos plasmodios.[17] [18] [19] [14]
Hongos
La mayoría de los hongos crecen como hifas, estructuras cilíndricas y filiformes de 2 a 10 micrómetros de diámetro y hasta varios centímetros de longitud. Las hifas nuevas se forman típicamente por la aparición de nuevos ápices a lo largo de hifas preexistentes por un proceso llamado de ramificación, aunque también puede ocurrir que el extremo apical de las hifas se bifurque, dando lugar a dos hifas con crecimiento paralelo.[20] Las hifas pueden ser septadas o cenocíticas: las primeras se hallan divididas en compartimentos con un sólo núcleo por medio de tabiques internos, o septos, que se forman en ángulo recto con las paredes de la hifa. Las hifas cenocíticas, en cambio, no están compartimentalizadas por lo que forman grandes células multinucleadas.[21]
En muchos hongos filamentosos inferiores el micelio está muy poco dividido, presentando escasos septos, por lo que forma verdaderos sincitios. En cambio, la mayoría de los hongos filamentosos superiores presentan dicarión, en el cual las células filiformes se hallan parcialmente divididas en segmentos, cada uno con dos núcleos diferentes,[22] provenientes de distintas células madres y que no se fusionan.[23]
Plantas superiores
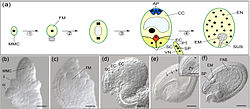 Desarrollo del gametofito femenino y del embrión en Arabidopsis. (a) Esquema que muestra la ontogenia del gametofito femenino y el desarrollo temprano del embrión y del endosperma. Una megaspora haploide funcional (FM) se desarrolla a partir de una célula madre de la megaspora (MMC) diploide a través de meiosis (1). Después de tres mitosis sin citocinesis, la FM se convierte en un sincitio con 8 núcleos (2). Luego de la migración de los núcleos dentro del sincitio, la formación de paredes celulares y la diferenciación (3), se desarrolla un saco embrionario con 7 células y 8 núcleos. Este saco contiene una oósfera (EC) y dos sinérgidas (SC) en el polo micropilar, tres antípodas (AP) en el polo chalazal y una célula central vacuolizada que lleva dos núcleos (CC). Las tres antípodas degeneran más tarde, y la disolución de un asinérgida precede a la entrada del tubo polínico (TP). Dos núcleos generativos (SP) se unen, independientemente, a la óosfera y a los núcleos polares, originando el embrión (EM) y el endosperma (EN), respectivamente, a través del proceso de doble fecundación. SUS: suspensor; VN: núcleo vegetativo del tubo polínico. (b-f) Micrografías de secciones transversales de óvulos correspondientes a los sucesos representativos descritos previamente. ii: tegumento interno; oi: tegumento externo del óvulo. Las divisiones mitóticas (mostradas con flechas en "e") del núcleo triploide formado luego de la fusión de un núcleo generativo con los dos núcleos polares lleva al desarrollo de un endosperma sincitial (FNE) como se muestra en "f".[24]
Desarrollo del gametofito femenino y del embrión en Arabidopsis. (a) Esquema que muestra la ontogenia del gametofito femenino y el desarrollo temprano del embrión y del endosperma. Una megaspora haploide funcional (FM) se desarrolla a partir de una célula madre de la megaspora (MMC) diploide a través de meiosis (1). Después de tres mitosis sin citocinesis, la FM se convierte en un sincitio con 8 núcleos (2). Luego de la migración de los núcleos dentro del sincitio, la formación de paredes celulares y la diferenciación (3), se desarrolla un saco embrionario con 7 células y 8 núcleos. Este saco contiene una oósfera (EC) y dos sinérgidas (SC) en el polo micropilar, tres antípodas (AP) en el polo chalazal y una célula central vacuolizada que lleva dos núcleos (CC). Las tres antípodas degeneran más tarde, y la disolución de un asinérgida precede a la entrada del tubo polínico (TP). Dos núcleos generativos (SP) se unen, independientemente, a la óosfera y a los núcleos polares, originando el embrión (EM) y el endosperma (EN), respectivamente, a través del proceso de doble fecundación. SUS: suspensor; VN: núcleo vegetativo del tubo polínico. (b-f) Micrografías de secciones transversales de óvulos correspondientes a los sucesos representativos descritos previamente. ii: tegumento interno; oi: tegumento externo del óvulo. Las divisiones mitóticas (mostradas con flechas en "e") del núcleo triploide formado luego de la fusión de un núcleo generativo con los dos núcleos polares lleva al desarrollo de un endosperma sincitial (FNE) como se muestra en "f".[24]En las gimnospermas, la megáspora funcional se divide por mitosis muchas veces, iniciando la formación del gametofito femenino. En el caso particular del pino hay una etapa inicial de divisiones nucleares que no son seguidas de la formación de paredes celulares, lo que origina un cenocito con unos 2.000 núcleos, estado en el que pasa el invierno. En la siguiente primavera reanuda el crecimiento formando paredes celulares entre los núcleos del cenocito, constituyendo el endosperma primario o prótalo. Cuando terminan de formarse las paredes celulares, se forman dos o tres arquegonios en el prótalo hacia el extremo donde se halla la micrópila. El prótalo y los arquegonios constituyen el gametofito femenino maduro.[25]
En el proceso de doble fecundación en angiospermas, uno de los núcleos generativos del grano de polen se fusiona con la oósfera para originar el cigoto, mientras que el otro núcleo generativo se une con los núcleos polares de la célula central del saco embrionario para formar el núcleo triploide de la célula central o célula a partir de la cual se desarrollará el endosperma. El mismo puede ser de tres tipos, dependiendo del tipo de proceso de formación. En el tipo celular la cariocinesis está acompañada con la citocinesis, por lo que únicamente se originan células uninucleadas. Por el contrario, en los tipos nuclear y helobial las primeras divisiones del núcleo de la célula endospermogenética no están acompañadas de la división del citoplasma a través de paredes celulares, por lo que se forman células cenocíticas que posteriormente pueden formar paredes celulares.[26] [27] [28] En el endosperma de los cereales la etapa cenocítica de desarrollo se caracteriza por divisiones celulares sincronizadas alrededor de una gran vacuola central siguiendo un patrón de polaridad antero-posterior. Esta etapa transcurre de 3 a 5 días luego de la doble fertilización, dependiendo de la especie y de las condiciones ambientales y es inmediatamente seguida por una serie de citocinesis que transforman al cenocito en un tejido multicelular.[29] [30]
Invertebrados
Poríferos
Algunas clases de poríferos como las esponjas vítreas (de las que se conocen unas 500 especies)[31] poseen un esqueleto mineral compuesto por espículas silíceas.[32] Sus cuerpos suelen tener forma de vaso y están formados tanto por tejidos celulares como sincitiales, hasta el punto en que en algunas especies la mayor parte de estos animales tienen naturaleza sincitial, incluyendo la capa de coanocitos y todos los tejidos en contacto con el agua.[33]
Ernst Haeckel estudió estas especies, desarrollando su teoría de la biocristalización o de formación de las espículas,[34] en la que consideraba que las espículas eran estructuras intermedias entre el carbonato de calcio y las células del sincitio; una especie de secreción que la selección natural había elegido para conformar el esqueleto de las esponjas. De esta forma el biólogo alemán acuñó el término sincitio en 1872,[35] aunque su teoría resultó ser incorrecta.[34]
Insectos
El sincitio en el embrión temprano de invertebrados es importante para la diferenciación celular. En el caso de Drosophila melanogaster por ejemplo, después de la fusión del espermatozoide con el oocito comienza un proceso de mitosis incompleta en el cual se dividen los núcleos de las células pero no hay segmentación del citoplasma (no hay formación de nuevas membranas celulares).[7] Las primeras divisiones ocurren en el centro del huevo. Cuando se han formado 256 núcleos, la mayoría migra a la periferia para seguir proliferando[37] hasta originar un sincitio de unos 6 000 núcleos al final de 12 divisiones sucesivas.[7] En torno a cada núcleo se encuentran proteínas y estructuras del citoesqueleto —como microtúbulos y microfilamentos— formando pequeñas islas conocidas como energidas. Debido a la presencia de éstas, el citoplasma en que están inmersos los núcleos no es homogéneo. Esta etapa es conocida como blastodermo sincitial, y corresponde al blastodermo o blástula en otras especies. En el ciclo 9 de división cinco núcleos migran hacia el polo posterior del huevo, diferenciándose en células polares que finalmente originarán nuevos gametos. A partir del decimotercer ciclo de división, los núcleos son separados por plegamientos hacia el interior de la membrana del huevo, para formar células individuales que constituirán el blastodermo celular, el cual se dispone como una sola capa en torno al vitelo.[37]
La formación de un sincitio en esta especie facilita la difusión de proteínas y otras moléculas de gran tamaño como el ARNm que codifica los factores de transcripción Bicoid y Nanos y otros morfogenes. La proteína Bicoid se expresa en un gradiente que va desde el extremo anterior del embrión temprano al posterior, mientras que la proteína Nanos se concentra en el extremo posterior del sincitio. Los núcleos con más Bicoid activan los genes que promueven la diferenciación de las células en la cabeza y las estructuras del tórax. Los núcleos expuestos a más Nanos activan los genes responsables de la diferenciación de las regiones posteriores, como el abdomen y las células germinales.[7]
La especificación del eje dorso-ventral se basa en el gradiente de transcripción del factor de transcripción Dorsal. Una mayor concentración de Dorsal en la parte ventral del huevo determina el destino ventral de esas células, mientras que su ausencia establece el dorso del animal. Sin embargo, a diferencia de Bicoid, Dorsal no establece su gradiente dentro del sincitio, sino sobre ciertas células señalizadas por procesos célula-célula.[37] Adicionalmente, los genes gap que dictarán la futura estructura segmentada del cuerpo, se manifiestan ya en esta etapa del desarrollo embrionario.[7]
Vertebrados
Músculo
- Músculo esquelético
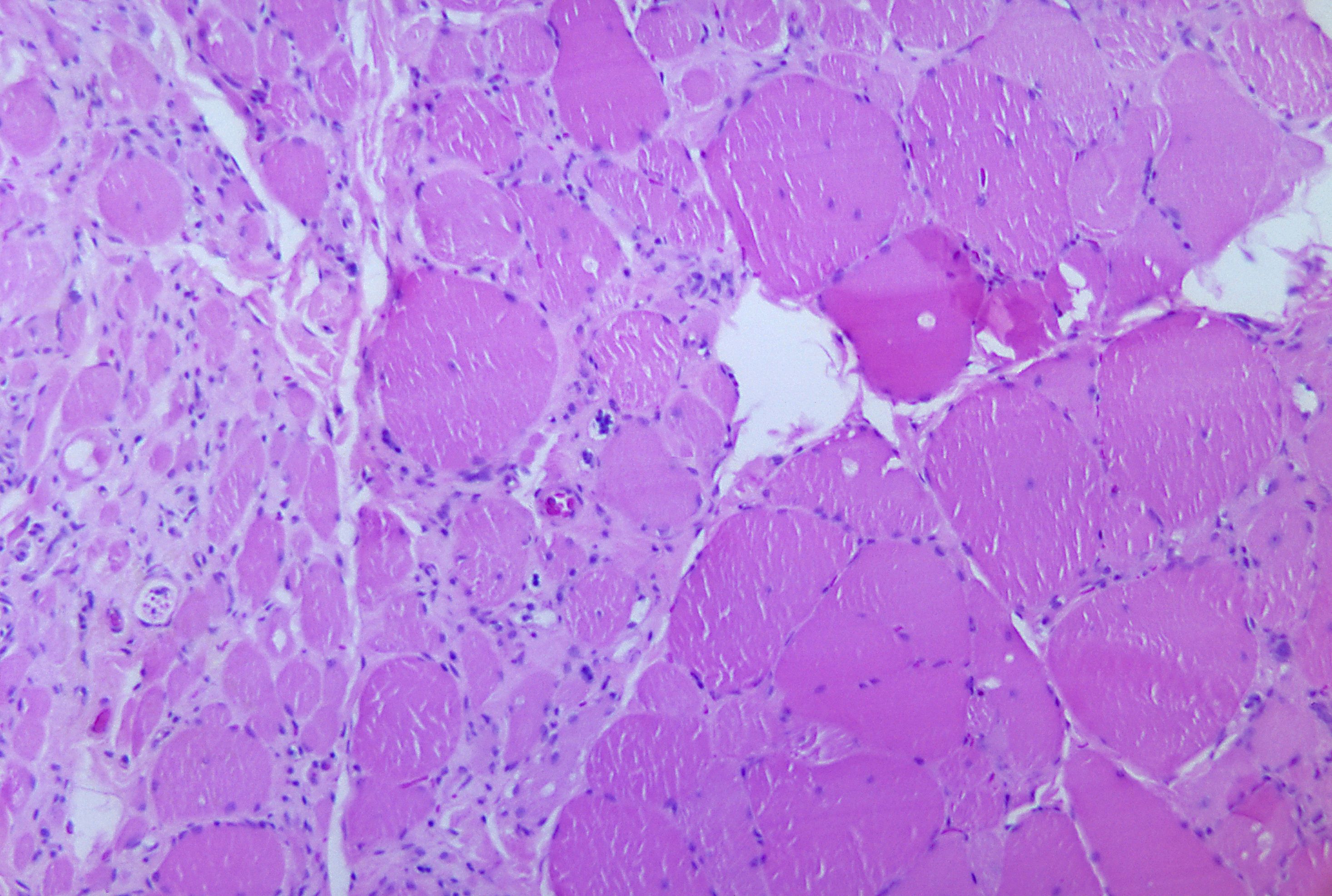
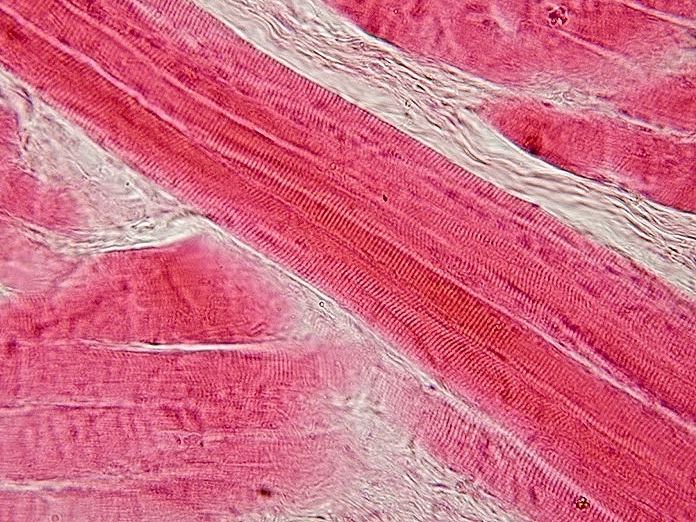
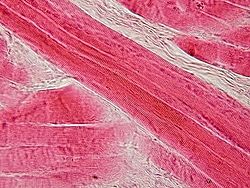 Corte longitudinal de músculo esquelético.
Corte longitudinal de músculo esquelético.El músculo esquelético es considerado un verdadero sincitio anatómico, pues cada célula presenta múltiples núcleos, originada por la fusión de múltiples células. Los mioblastos comienzan a multiplicarse, influidos por factores de crecimiento como el FGF. Los receptores integrinas de las fibronectinas y otras moléculas como las cadherinas los obligan a reconocerse, alinearse y adherirse, para posteriormente fusionarse en células musculares sincitiales. Este último proceso está mediado por meltrinas (unas metaloproteasas).[9]
El sincitio del músculo esquelético permite una rápida contracción coordinada de los músculos en toda su longitud. El potencial de acción se propaga a lo largo de la superficie de la fibra muscular desde el punto de contacto sináptico con la neurona motora.[38] En estados patológicos como la miopatía, la presencia de sincitio permite la viabilidad muscular, porque los focos de necrosis de una parte del músculo esquelético no resultan en la necrosis de las secciones adyacentes de esa zona ya que estas regiones tienen su propio material nuclear, aunque algunos de los segmentos supervivientes queden sin inervación por la pérdida de continuidad con la unión neuromuscular.[39]
- Músculo cardíaco
Al igual que el músculo esquelético, el músculo cardíaco también es un músculo con estriaciones debido a la presencia de retículo sarcoplásmico y miofibrillas. Inicialmente los investigadores consideraban que la fibra muscular cardíaca era un sincitio como la fibra esquelética. Sin embargo, con el desarrollo de la microscopía electrónica pudo comprobarse que no es un auténtico sincitio anatómico,[40] porque cada fibra miocárdica se encuentra separada de las vecinas por sus respectivos sarcolemas en la zona lateral, que se continúan con los discos intercalares en el extremo de cada fibra.[38]
Estos discos tienen uniones en hendidura, con conductancias muy elevadas,[38] que conectan los citoplasmas de células vecinas lo que lleva a que cuando se recibe un potencial de acción se produzca una onda de despolarización (precediendo a la contracción de las aurículas y de los ventrículos) sin que exista un neurotransmisor que desencadene la respuesta,[41] por lo que, si bien el músculo cardíaco no es un sincitio anatómico, se comporta como un sincitio funcional. Los discos intercalares además podrían influir en la cohesión del tejido cardíaco durante la contracción.[40] En el músculo cardíaco, el proceso de formación de las células del tejido es similar al de D. melanogaster (por división, no fusión), finalizando con la expresión de miogenina, una proteína que comienza el proceso para que se produzca la diferenciación final de la célula muscular.[9]
- Músculo liso
A diferencia de los tejidos esqueléticos estriados, el músculo liso contiene retículos escasamente desarrollados y dispersos por la célula, que se contraen debido a las variaciones de la concentración de iones de calcio. El músculo liso puede tener una sola o múltiples unidades. Si bien ninguno de ellos son sincitios anatómicos, el músculo liso de unidad única o visceral puede actuar como un sincitio eléctrico (funcional) debido a que las células están relacionadas por uniones en hendidura, que permiten el pasaje del impulso nervioso y la contracción coordinada como respuesta al estímulo producido por un potencial de acción. Este tejido es abundante en el abdomen y en órganos del aparato digestivo como el intestino.[41] Allí, la presencia de un sincitio funcional en el músculo liso que conforman sus paredes produce que al ser excitado se forme un anillo contráctil que expande el estímulo hacia zonas contiguas adelante y atrás del punto en que se recibió.[42] Por el contrario, el de unidades múltiples o multiunitario tiene igualmente contracciones involuntarias pero en forma independiente debido a que las células están aisladas eléctricamente y por lo tanto no es un sincitio en ningún aspecto. Este tejido se puede encontrar por ejemplo en el iris de los ojos.[38] [41]
Placenta
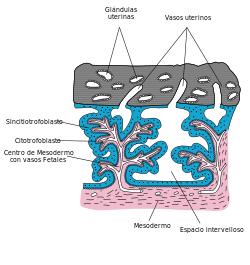 Esquema de un corte transversal de placenta, mostrando el citotrofoblasto, el sincitiotrofoblasto y otras estructuras, especialmente los vasos sanguíneos.
Esquema de un corte transversal de placenta, mostrando el citotrofoblasto, el sincitiotrofoblasto y otras estructuras, especialmente los vasos sanguíneos.Otro sincitio importante en vertebrados está en la placenta de los mamíferos placentarios. La conversión del citotrofoblasto (capa de Langhans) en sincitio es una forma fisiológica para regenerar la placenta. Cuando se implanta el embrión está rodeado únicamente de elementos citotrofoblásticos. Sin embargo, algunos estudios científicos[43] [44] demostraron que en el día 14 después de la ovulación, cuando debería producirse la menstruación en condiciones de no - embarazo, la hemorragia periovular que ocurre induce la transformación en sincitio del trofoblasto inmaduro, debido a reacciones inmunitarias.[45]
Cuando se forma la placenta, el estrato de Langhans predomina sobre el sincitio. A medida que la placenta madura, la relación se invierte. La distribución característica de la circulación de la sangre por los vasos fetoplacentarios determinan una mejor irrigación tanto del estrato de Langhans como del sincitio, facilitando el intercambio entre la circulación fetal y el espacio intervelloso, lo que constituye una adaptación a la mejor oxigenación del feto en el embarazo.[46]
Las células embrionarias, sin membranas que las separen, forman una barrera multinucleada.[45] [46] El sincitio placentario tiene además otras funciones. Presenta gran cantidad de organelos citoplasmáticos, especialmente en las últimas etapas de desarrollo fetal, indicando un alto metabolismo. También es un gran depósito de hierro, que obtiene del plasma materno.[47] Además se ha demostrado mediante estudios inmunohistoquímicos que el sincitio es el principal lugar donde se forma la gonadotrofina coriónica humana (hCG). Así mismo, como se ha detectado en el huevo recién implantado en el cual el trofoblasto aún no ha madurado, se piensa que también se origina en el citotrofoblasto. El sincitio produce gran cantidad de proteínas placentarias que vierte a la sangre materna. Algunos afirman que evitan el rechazo del feto por procesos inmunes naturales y evitan la coagulación sanguínea en los espacios intervellosos, aunque otros piensan que no cumplen ninguna función. Estas proteínas son liberadas por factores liberadores producidos por la propia placenta, que de este modo actúa en forma autócrina. La producción de proteínas se mantiene en permanente aumento durante todo el embarazo.[45]
Cuando el embarazo acaba, el citotrofoblasto se disgrega y el sincitio se vuelve extremadamente delgado, produciendo un estrecho acercamiento entre la sangre materna y la fetal, hasta el punto que en algunos sitios quedan apenas separadas por el endotelio de los capilares fetales. Estos puntos se conocen como membranas vásculo-sincitiales, que se ubican por debajo del sincitio y lo separan del capilar fetal (semejante a la del glomérulo renal). Estas membranas están especializadas en filtrar la sangre; no se encuentran en placentas inmaduras, ni en maduras patógenas en las que el trofoblasto no ha madurado. Puede considerarse entonces que la placenta se divide en dos superficies con funciones distintas: la que corresponde a la aposición endotelio-corial (filtrante) y otra constituida por el sincitio (metabólica).[45]
Sistema nervioso central
Las primeras teorías sobre el sistema nervioso central, desarrolladas a partir de la segunda mitad del siglo XIX, planteaban que las células se encontraban unidas[48] formando un sincitio,[49] y que las terminaciones nerviosas se entrelazaban entre sí formando una red. Esta teoría, refinada por Joseph von Gerlach, fue llamada reticularismo o teoría reticular, y permitía entre otras cosas, explicar la transmisión del impulso eléctrico entre neuronas. His y Forel se opusieron a esta idea, pero carecían de metología científica que los avalara.[50] En 1872 Camilo Golgi desarrolló un método de tinción para analizar al microscopio preparados histológicos de tejido nervioso,[51] que hoy se conoce como tinción de Golgi. La técnica, basada en la precipitación de sales de plata, producía siluetas negras de las células, pero sus resultados no sólo eran inconsistentes sino que ocultaban la estructura celular y las terminaciones nerviosas, pudiendo llevar a interpretaciones erróneas de los mismos. Posteriormente Santiago Ramón y Cajal refinaría esta técnica desarrollando una propia, llamada "de plata reducida" o tinción de Cajal, que permitía visualizar la morfología neuronal. Aplicando estas técnicas a tejidos de embriones y animales jóvenes, Cajal no solo probó que no existía un sincitio en el sistema nervioso central, sino que desarrolló la teoría moderna de la neurona, explicando además el crecimiento de las prolongaciones nerviosas e incursionando en el estudio de la polaridad del impulso nervioso.[52]
A pesar de ello, modernamente se ha establecido que algunas células que componen el sistema nervioso como los astrocitos o algunos axones gigantes pueden ser sincitiales. Los astrocitos aíslan las sinapsis de otras terminaciones nerviosas adyacentes y del medio extracelular. Los astrocitos presentan las mismas uniones en hendidura que el músculo cardíaco, quedando unidos formando un sincitio. Esto permite el intercambio de iones y otras moléculas de pequeño tamaño (como los neurotransmisores), haciendo que los astrocitos actúen a modo de buffer,[53] en especial de la concentración de ion potasio en el sistema nervioso.[54] Esto es importante en algunos casos como la hipoxia, en que la concentración local de este ion puede aumentar hasta 20 veces, despolarizando las membranas y provocando la liberación de neurotransmisores que multipliquen el efecto, pudiendo llevar a la muerte neuronal.[53]
Los axones pueden estar formados por una o muchas células. Dentro de los axones multicelulares se encuentran los axones gigantes con mielina, que pueden clasificarse en segmentados o sincitiales. Un ejemplo de axón sincitial está representado por los axones gigantes de calamar, que se obtienen por fusión de unas 300 a 500 células.[55]
Sincitios formados por la interacción hospedador-patógeno
Nematodos
Los nematodos parásitos, tanto de plantas como de animales, inducen la fusión de las células del hospedero formando un sincitio sin el cual no podrían alimentarse, desarrollarse y completar su ciclo vital. Globodera rostochiensis y G. pallida, conocidos como los «nematodos del quiste de la papa», son nematodos endoparásitos sedentarios de plantas pertenecientes a la familia de las solanáceas, como la papa y el tomate, y se consideran una de las plagas más importante de estos cultivos.[56]
Las raíces en crecimiento activo de las plantas susceptibles al parásito exudan sustancias estimulantes, entre las que se incluye el glicoalcaloide solanina que induce los cambios fisiológicos como modificaciones en la estructura de las glándulas esofágicas y en la expresión génica. Ambas especies de nematodos reaccionan a estos estimulantes eclosionando como juveniles infectivos que penetran en la zona de elongación de las raíces y migran a través de las células hasta la endodermis que rodea el cilindro vascular. El nematodo introduce productos en las células corticales adyacentes a la endodermis que rodean la cabeza del nematodo induciéndoles una profunda transformación. De tres a diez de esas células se funden entre sí, sus paredes celulares se engrosan, el citoplasma se torna denso y se origina un sincitio con una alta actividad metabólica, el cual es indispensable para que el parásito pueda completar su ciclo de vida pues le proporciona los nutrientes necesarios para su crecimiento y desarrollo.[57] [58] El sincitio presenta una alta actividad de ribosomas y un retículo endoplásmico rugoso dispuesto en forma concéntrica entre los que se incluyen otros organelos como mitocondrias y plastidios distribuidos en el citoplasma, lo que denota síntesis metabólica. El sincitio también es utilizado por las hembras del parásito para poner huevos. Por el contrario, en cultivos resistentes al nematodo, aunque las primeras etapas de la infección se desarrollan en forma similar, la papa produce una reacción interna que forma un tejido similar al sincitio pero limitado por una membrana que no permite que los parásitos juveniles puedan madurar.[59]
La triquinelosis es una enfermedad parasitaria distribuida a nivel mundial que afecta a mamíferos silvestres y domésticos y que se transmite de modo accidental a los seres humanos por ingestión de carne o productos cárnicos crudos o insuficientemente cocidos procedentes de animales infectados.[60] Esta enfermedad es causada por diferentes especies, subespecies y cepas de nematodos del género Trichinella. La infección se inicia después de ingerir un quiste, que en su interior presenta una larva enrollada en forma de espiral. Al digerir el quiste en el estómago, las larvas son liberadas y transportadas pasivamente por peristaltismo al yeyuno donde se desarrolla el parásito adulto. Las hembras del parásito perforan la mucosa intestinal formando un sincitio por fusión de aproximadamente 120 células intestinales. Las hembras se alimentan y desarrollan en su interior, produciendo larvas que se distribuyen por todo el organismo hospedero por vía linfo-hemática para terminar enquistándose en su musculatura estriada. El sincitio les sirve además como protección.[61]
Virus
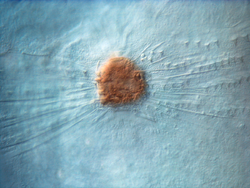
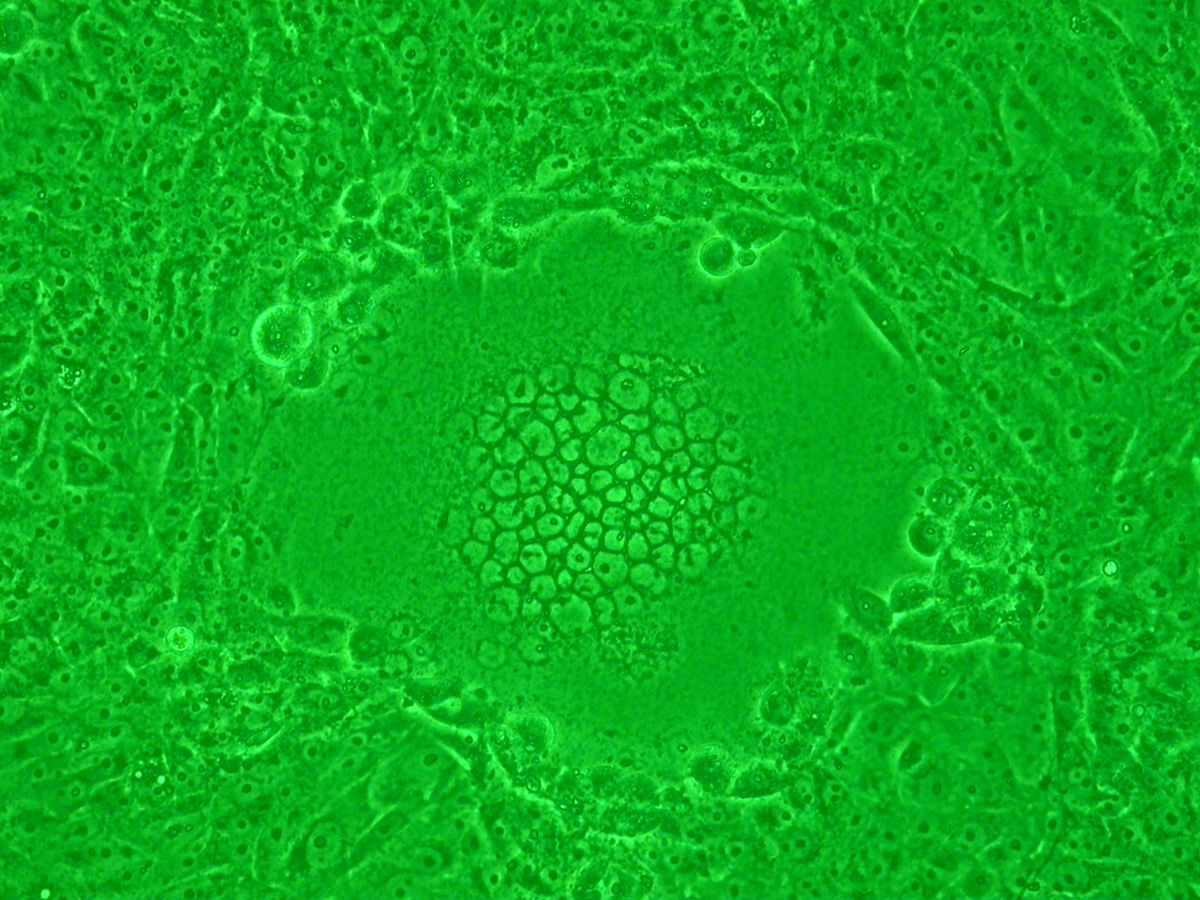
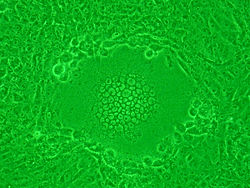 Sincitio causado por la infección HSV-1 en células Vero.
Sincitio causado por la infección HSV-1 en células Vero.Ciertos tipos de virus como los de la familia Paramyxoviridae (incluyendo el virus sincitial respiratorio (RSV)[62] y el VIH también forman sincitios, pero con las células del huésped.[63] En la infección por el VIH, el virus puede infectar un linfocito T cooperador. Entonces, la célula comienza a mostrar las glicoproteínas superficiales del VIH, que son antígenoicas. Normalmente, un linfocito T citotóxico inmediatamente llega a "inyectar" linfotoxinas, como perforina o granzima, que matan a la célula T ayudante infectada. Sin embargo, si hay cerca células T colaboradoras, los receptores gp41 del VIH que aparecen en la superficie de la célula T se une a otros linfocitos similares. Esto hace que decenas de células T colaboradoras fusionen sus membranas celulares en un sincitio gigante no funcional, lo que hace que el virión del VIH pueda matar muchas células T colaboradoras mediante la infección de una sola.[64]
El virus sincitial respiratorio bovino (VSRB) obliga a que las células no ciliadas del epitelio de los bronquios y alvéolos proliferen formando células sincitiales de gran tamaño, con acumulación de linfocitos en el tejido intersticial.[65] En al menos la mitad de los lactantes internados por bronquiolitis en Estados Unidos el agente etiológico es virus sincitial respiratorio. El virus induce la formación de un sincitio en las vías aéreas de los infantes que probablemente provoca una reacción a la inmunoglobulina E y a sustancias liberadas por los neutrófilos, produciendo edema en la mucosa, broncoespasmo y finalmente la necrosis del epitelio del sistema respiratorio debido a que las células gigantes del sincitio ocluyen los alvéolos, los bronquiolos y los bronquios, dificultando la respiración.[66]
Véase también
Referencias
- ↑ Equipo Staff (1998). «24 "S"». Diccionario médico (4ª, reimpresa edición). Elsevier España. p. 629. ISBN 9788445804865. http://books.google.com.uy/books?id=OmPwLAv65C4C&pg=PA629&dq=sincitio&hl=es&ei=kIAlTqPzMsrl0QHL5IzrCg&sa=X&oi=book_result&ct=result&resnum=8&ved=0CFEQ6AEwBw#v=onepage&q=sincitio&f=false.
- ↑ García Peralta, Faustino (1973). Minerva Books. ed. Fundamentos de biología (10ª edición). p. 72. http://books.google.com.uy/books?id=kaBcAAAAMAAJ&q=sincicio&dq=sincicio&hl=es&ei=sXclTue3Kefx0gGnq9TFCg&sa=X&oi=book_result&ct=result&resnum=9&ved=0CEsQ6AEwCA.
- ↑ González, A.M. & Arbo, M.M.. «Glosario». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de febrero de 2011.
- ↑ El término se aplica a especies con pared celular. Dorland (2005). Dorland Diccionario enciclopédico ilustrado de Medicina (30ª edición). Elsevier España. p. 344. ISBN 9788481747904. http://books.google.com.uy/books?id=E3u7aczu1_cC&pg=PA344&dq=cenocito&hl=es&ei=CKg4TrbqLeXn0QGsye3pAw&sa=X&oi=book_result&ct=result&resnum=7&ved=0CEYQ6AEwBg#v=onepage&q=cenocito&f=false.
- ↑ Watson, James (2006). Biología molecular del gen (5ª edición). Ed. Médica Panamericana. p. 631. ISBN 9788479035051. http://books.google.com.uy/books?id=ajwHxb6peRkC&pg=PA631&dq=sincitio+drosophila&hl=es&ei=sNDbTaPOK6b20gGdvq3lDw&sa=X&oi=book_result&ct=result&resnum=4&ved=0CDUQ6AEwAw#v=onepage&q=sincitio%20drosophila&f=false.
- ↑ Muñoz Martínez, E. J.; García, X.. «8 Citoesqueleto». En Fondo de Cultura Económica. Fisiología I. Células, órganos y sistemas. p. 78. ISBN 9789681654467. http://books.google.com.uy/books?id=tX1XSZuGOfcC&pg=PA78&dq=la+mayor%C3%ADa+de+las+c%C3%A9lulas+tienen+un+solo+n%C3%BAcleo&hl=es&ei=-dwoTtuPBe-z0AHbhojPCg&sa=X&oi=book_result&ct=result&resnum=7&ved=0CEYQ6AEwBjgK#v=onepage&q=la%20mayor%C3%ADa%20de%20las%20c%C3%A9lulas%20tienen%20un%20solo%20n%C3%BAcleo&f=false.
- ↑ a b c d e f g Wolpert, Lewis (2010). «2: Ciclo vital y desarrollo general de Drosophila». En Ed. Médica Panamericana. Principios Del Desarrollo / Development Principles (3ª edición). pp. 33-84. ISBN 9788498352061. http://books.google.com.uy/books?id=HCkurx8FuSgC&pg=PA33&dq=sincitio+drosophila&hl=es&ei=sNDbTaPOK6b20gGdvq3lDw&sa=X&oi=book_result&ct=result&resnum=1&ved=0CCUQ6AEwAA#v=onepage&q=sincitio%20drosophila&f=false.
- ↑ Mendivil Navarro, Javier. «Biología General. La Célula: Citología. Los seres pluricelulares.». Asociación Cultural Aragón Interactivo y Multimedia. Consultado el 11 de julio de 2011.
- ↑ a b c Eynard, Aldo; Valentich, Mirta; Rovasio, Roberto (2008). «Parte II - Interacciones celulares, patrones complejos e hostogénesis». En Ed. Médica Panamericana. Histología y Embriología del Ser Humano/ Histology and Embryology of the Human Being: Bases Celulares Y Moleculares/ Cellular and Molecular Basis (4ª edición). pp. 264-265. ISBN 9789500606028. http://books.google.com.uy/books?id=p1JSyuGai0oC&pg=PA264&dq=formaci%C3%B3n+c%C3%A9lula+muscular+esquel%C3%A9tica&hl=es&ei=3erbTcDVAuj20gG_9JDBDw&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCoQ6AEwAQ#v=onepage&q=formaci%C3%B3n%20c%C3%A9lula%20muscular%20esquel%C3%A9tica&f=false. Consultado el 24 de mayo de 2011.
- ↑ «Diccionario médico». Médicos cubanos.com (Setiembre de 2008). Consultado el 30 de julio de 2011.
- ↑ Ross, Michael H.; Wojciech, Pawlina (2007). Histología (5ª edición). Ed. Médica Panamericana. p. 321. ISBN 9789500604352. http://books.google.com.uy/books?id=NxYmIRZQi2oC&pg=PA321&dq=diferenciaci%C3%B3n+de+mioblasto&hl=es&ei=oU3dTYz-Gonu0gGK-fXwDw&sa=X&oi=book_result&ct=result&resnum=5&ved=0CDwQ6AEwBDgK#v=onepage&q&f=false.
- ↑ Jacobs, W. P. (1994). «Caulerpa» (en inglés). Scientific American 271 (6): pp. 66–71. ISSN 0036-8733.
- ↑ Jensen, M. N.. «Caulerpa, The World's Largest Single-celled Organism?» (en inglés). Consultado el 29 de mayo de 2011.
- ↑ a b Bachmann, Konrad (1978). Biología para médicos: conceptos básicos para las facultades de medicina, farmacia y biología.. Volumen 2 de Serie de Biología Fundamental. Reverte. p. 196. ISBN 9788429118049. http://books.google.com.uy/books?id=dwPg88Wq2xYC&pg=PA196&dq=sincitio+algas&hl=es&ei=QxPdTaXjPOrm0QH2ztjdDw&sa=X&oi=book_result&ct=result&resnum=1&ved=0CCwQ6AEwAA#v=onepage&q&f=false. Consultado el 25 de mayo de 2011.
- ↑ Naturaleza Educativa. «Botánica. Clasificación. Ficofitos o algas - 2ª parte». Asociación Española para la Cultura, el Arte y la Educación (ASOCAE O.N.G.D.). Consultado el 1 de agosto de 2011.
- ↑ Des Abbayes, H. (1989). Botánica: vegetales inferiores. Reverte. pp. 283 - 285. ISBN 9788429118131. http://books.google.com.uy/books?id=zjqqXtzghvoC&pg=PA283&dq=algas+sifonocladales&hl=es&ei=cAY2TvaaBtO70AGAyOjyCw&sa=X&oi=book_result&ct=result&resnum=3&ved=0CDEQ6AEwAg#v=onepage&q&f=false. Consultado el 1 de agosto de 2011.
- ↑ Baldauf SL, Doolittle WF (octubre 1997). «Origin and evolution of the slime molds (Mycetozoa)» (en inglés). Proc. Natl. Acad. Sci. U.S.A. 94 (22): pp. 12007–12. doi:. PMID 9342353. PMC 23686. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=9342353.
- ↑ Fiore-Donno AM, Berney C, Pawlowski J, Baldauf SL (2005). «Higher-order phylogeny of plasmodial slime molds (Myxogastria) based on elongation factor 1-A and small subunit rRNA gene sequences» (en inglés). J. Eukaryot. Microbiol. 52 (3): pp. 201–10. doi:. PMID 15926995.
- ↑ «Arbor». Consejo Superior de Investigaciones Científicas (España) 34. 1956. http://books.google.com.uy/books?id=hhIRAAAAIAAJ&q=hongo+sincitio&dq=hongo+sincitio&hl=es&ei=2v7cTZfLMKHn0QG6gOm_Dw&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCkQ6AEwATgU.
- ↑ Harris SD. (2008). «Branching of fungal hyphae: regulation, mechanisms and comparison with other branching systems» (en inglés). Mycologia 50 (6): pp. 823–32. doi:. PMID 19202837.
- ↑ Chang S-T, Miles PG. (2004) (en inglés). Mushrooms: Cultivation, Nutritional Value, Medicinal Effect and Environmental Impact. CRC Press. ISBN 0849310431.
- ↑ Prats Pastor, Guillermo (2006). «5 - Micología». Microbiología Clínica. Ed. Médica Panamericana. pp. 83-84. ISBN 9788479039714. http://books.google.com.uy/books?id=TdsoWPEYaoUC&pg=PA83&dq=hongo+sincitio&hl=es&ei=_vPcTczCFuXV0QHk0NQE&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCoQ6AEwAQ#v=onepage&q&f=false.
- ↑ Piaggio, Mario (Última renovación 9 de julio de 2008). «Curso de biología vegetal. Atlas de imágenes. "D"». Sección Micología de la Facultad de Ciencias (UDELAR). Consultado el 2 de agosto de 2011.
- ↑ Johnston, A. J.; Meier, P.; Gheyselinck, J.; Wuest, S.E.J.; Federer, M.; Schlagenhauf, E.; Becker, J.D.; Grossniklaus, U. (2007). «Genetic subtraction profiling identifies genes essential for Arabidopsis reproduction and reveals interaction between the female gametophyte and the maternal sporophyte» (en inglés). Genome Biology 8 (10): pp. R204-R204.21. http://genomebiology.com/2007/8/10/R204.
- ↑ González, A.M. & Arbo, M.M.. «Megasporogénesis y megagametogénesis en pino». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de mayo de 2011.
- ↑ Vijayaraghavan, M.R.; Prabhakar, K. (1984). «The endosperm» (en inglés). Johri B.M., ed. Embryology of Angiosperms. Berlin (Springer-Verlag): pp. 319–376.
- ↑ Lopes, M.A.; Larkins, B.A. (1993). «Endosperm origin, development, and function» (en inglés). The Plant Cell Online 5 (10): pp. 1383-1399. ISSN 1532-298X. http://www.plantcell.org/content/5/10/1383.full.pdf+html?ijkey=95ca5db4507d780e90f0af309448cd7f1aef1df4&keytype2=tf_ipsecsha.
- ↑ Berger, F. (1999). «Endosperm development» (en inglés). Curr. Opin. Plant Biol. 2: pp. 28-32. ISSN 1369-5266.
- ↑ Berger, F. (2001). «Dynamic Analyses of the Expression of the HISTONE::YFP Fusion Protein in Arabidopsis Show That Syncytial Endosperm Is Divided in Mitotic Domains» (en inglés). Plant Cell 13: pp. 495-509. ISSN doi:10.1105/tpc.13.3.495. http://www.plantcell.org/content/13/3/495.full#R18.
- ↑ Kranz, E.; von Wiegen, P.; Quader, H.; Lörz, H. (1998). «Endosperm Development after Fusion of Isolated, Single Maize Sperm and Central Cells in Vitro» (en inglés). Plant Cell 10: pp. 511-524. ISSN doi:10.1105/tpc.10.4.511. http://www.plantcell.org/content/10/4/511.full.
- ↑ Museum of Paleontology. «Porifera - Hexactinellida: Life History and Ecology» (en inglés). University of California. Consultado el 10 de julio de 2011.
- ↑ Museum of Paleontology. «Porifera - Hexactinellida» (en inglés). University of California. Consultado el 10 de julio de 2011.
- ↑ Museum of Paleontology. «Porifera - Hexactinellida: More on Morpholgy» (en inglés). University of California. Consultado el 10 de julio de 2011.
- ↑ a b «Boletín». Sociedad Española de Historia Natural 5-6: pp. 374. 1905. http://books.google.com.uy/books?id=g7AsAQAAIAAJ.
- ↑ Die Kalkschwümme I (377): pp. 482-483. 1872. En Thompson, D'Arcy. «5: Sobre las espículas y los esqueletos espiculares». En J. T. Bonner. Sobre el crecimiento y la forma. Biología y diseño. Ediciones AKAL. pp. 149. ISBN 9788483233566. http://books.google.com.uy/books?id=rjesr-wC5CwC&pg=PA149&lpg=PA149&dq=Haeckel+sincitio&source=bl&ots=0lRO8eAFhb&sig=COakCwxw1uyQFmPO7zmxgw2UX8Y&hl=es&ei=aC7dTZuiKqLq0gGzzdHRDw&sa=X&oi=book_result&ct=result&resnum=5&ved=0CDMQ6AEwBA#v=onepage&q=Haeckel%20sincitio&f=false. Consultado el 31 de julio de 2011.
- ↑ Public Library of Science (2004). «The Molecular Biology of Wound Healing» (en inglés). PLoS Biology 2 (8). http://dx.doi.org/10.1371/journal.pbio.0020278.
- ↑ a b c Gilbert, S.F. (2005). «9 - La genética de especificación del eje en Drosophila». Biología del desarrollo con CD-Rom (7 edición). Médica Panamericana. pp. 286-287. ISBN 950-06-0869-3. http://books.google.com.uy/books?id=F6se5w-Z6uAC&printsec=frontcover&dq=Scott+Gilbert&hl=es&ei=CvlaTrKFI5TTgQe6w5yMDA&sa=X&oi=book_result&ct=result&resnum=1&ved=0CCwQ6AEwAA#v=onepage&q&f=false. Consultado el 29 de agosto de 2011.
- ↑ a b c d Levy, Matthew N.; Koeppen, Bruce M.; Stanton, Bruce A. (2006). Matthew N. Levy, Bruce M. Koeppen, Bruce A. Stanton. ed. Berne Y Levy Fisiologia (4ª edición). Elsevier España. ISBN 9788481749489.
- ↑ Stevens, Alan; Lowe, James (2001). «468». En Elsevier España. Anatomía Patológica (2ª edición). ISBN 9788481745122. http://books.google.com.uy/books?id=YkfRsm2Zl4AC&pg=PA468&dq=miopat%C3%ADa+necrosis&hl=es&ei=S_XbTbZwhpayA8_foLUO&sa=X&oi=book_result&ct=result&resnum=1&ved=0CCwQ6AEwAA#v=onepage&q=miopat%C3%ADa%20necrosis&f=false.
- ↑ a b Montoya Torno, Mario; Corporación para Investigaciones Biológicas (2002). «1». En Corporación para Investigaciones Biológicas. Cardiología (6ª edición). pp. 1-2. ISBN 9789589400579. http://books.google.com.uy/books?id=0-eN5A9Xk5YC&pg=PP17&dq=sincitio+m%C3%BAsculo+estriado&hl=es&ei=0j3LTa2FAcuDtge1o8z_Bw&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCoQ6AEwAQ#v=onepage&q&f=false.
- ↑ a b c Cunningham, James G. (2009). Elsevier España. ed. Fisiologia Veterinaria: Incluye Evolve. p. 87. ISBN 9788480863919.
- ↑ Montenegro, R. (2001). «6 - Sistema de órganos». En Raúl Montenegro. Biología evolutiva. Editorial Brujas. pp. 117-118. ISBN 9789874331816. http://books.google.com.uy/books?id=WA56vndSXNIC&pg=PA113&dq=frecuencia+sincitio&hl=es&ei=2JQ5ToGsBOfq0gG6mO3SAw&sa=X&oi=book_result&ct=result&resnum=6&ved=0CEIQ6AEwBTge#v=onepage&q=frecuencia%20sincitio&f=false. Consultado el 3 de agosto de 2011.
- ↑ Tuttle, SE; O´Toole, RW; O'Saughnessi, RW; Zuspan, FP (1985). «Immunohistochemical evaluation of human placental implantation: an initial study» (en inglés). Am J Obstet Gynecol 153 (3): pp. 239-44. http://www.ncbi.nlm.nih.gov/pubmed/3901762.
- ↑ Ulloa-Aguirre, A; Craviotto, A; Méndez, JP (1989). «Studies of the hCG secreted by human trophoblast in vitro» (en inglés). Proceed 71 Meeting Endocrine Society Seattle: pp. Abst # 501.
- ↑ a b c d Botella Llusiá, José (1993). Ediciones Díaz de Santos. ed. La placenta: fisiología y patología. p. 392. ISBN 9788479780654. http://books.google.com.uy/books?id=OYCa1Jq4BuYC&lpg=PA19&dq=sincitio%20placenta&pg=PA19#v=onepage&q=sincitio%20placenta&f=false.
- ↑ a b Botella Llusiá, José; Claveo Núñez, José A.. Tratado de ginecología: fisiología, obstetricia, perinatología, ginecología, reproducción (14ª edición). Ediciones Díaz de Santos. p. 101. ISBN 9788479780920. http://books.google.com.uy/books?id=CYdfAhqjG1IC&lpg=PA101&ots=p5HBUz1aah&dq=sincitio&pg=PA101#v=onepage&q=sincitio&f=false.
- ↑ Birkenfeld, A.; Mordel, N.; Okon, E. (1989). «Direct demonstration of iron in a term placenta in a case of beta-thalassemia major». Am. J. Obstet. Ginecol. 160 (3): pp. 562-563. ISSN 0002-9378. http://www.biomedsearch.com/nih/Direct-demonstration-iron-in-term/2929673.html.
- ↑ Outes, D.. Universidad de Morón (ed.): «En el centenario de la teoría de la neurona. Breve historia de como se relacionan las neuronas entre sí». Artes Gráficas Negri S.R.L.. Consultado el 3 de agosto de 2011.
- ↑ Gil-Loyzaga, P. E.. Cultivo de células animales y humanas. Editorial Visión Libros. p. 25. ISBN 9788499837376. http://books.google.com.uy/books?id=Z3t4R39FByYC&pg=PA25&lpg=PA25&dq=sincitio+reticularismo&source=bl&ots=qiRGKFmEMI&sig=Z_fITnQiVqjX2fzpmpE4NFqTT24&hl=es&ei=wcs4Tov_J6vC0AGz2d33Aw&sa=X&oi=book_result&ct=result&resnum=1&ved=0CBYQ6AEwAA#v=onepage&q=sincitio%20reticularismo&f=false.
- ↑ Ramón y Cajal, S.; Markram, H. (2007). Javier DeFelipe, Jorge Wagensberg. ed. Paisajes neuronales: homenaje a Santiago Ramón y Cajal. CSIC. pp. 49-51. ISBN 9788400085339. http://books.google.com.uy/books?id=jH5EEyeGliYC&pg=PA50&lpg=PA50&dq=teor%C3%ADa+++gerlach&source=bl&ots=xG_DnKyYIG&sig=um1EjpIuf6a4uee_NOXDZWaEwDU&hl=es&ei=0sE4Tq--Ecfk0QGswZS1Aw&sa=X&oi=book_result&ct=result&resnum=3&ved=0CCUQ6AEwAg#v=onepage&q=teor%C3%ADa%20%20%20gerlach&f=false.
- ↑ Medina Malo, Carlos (2004). Epilepsia: aspectos clínicos y psicosociales. Ed. Médica Panamericana. p. 33. ISBN 9789589181782. http://books.google.com.uy/books?id=ocSNl4uMiRYC&pg=PA33&dq=sincitio+golgi&hl=es&ei=MmblTeGnJcK5tgfX0KSDBw&sa=X&oi=book_result&ct=result&resnum=4&ved=0CDQQ6AEwAw#v=onepage&q=sincitio%20golgi&f=false.
- ↑ Estable, C. (1944). Ramón y Cajal - Homenaje en el 10º aniversario de su muerte (1934 - octubre 17 - 1944) (1000 ejemplares fuera de comercio edición). A92:61 RIO, Facultad de Química (UDELAR): Institución Cultural Española del Uruguay. pp. 29-36.
- ↑ a b Levy, Matthew N.; Berne, Robert M. (2009). Bruce A. Stanton, Bruce M. Koeppen. ed. Fisiología (6ª edición). Elsevier España. p. 56. ISBN 9788480864343. http://books.google.com.uy/books?id=lCSA7uxha0kC&pg=PA56&dq=sincitio+Schwann&hl=es&ei=-vziTfrYLaLg0QHCwpmpBw&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCsQ6AEwAQ#v=onepage&q=sincitio%20Schwann&f=false.
- ↑ Best (2010). «18 - Circulación coronaria e isquemia miocárdica». Bases fisiológicas de la práctica médica/ Physiological basis of medical practice (14ª edición). Ed. Médica Panamericana. p. 864. ISBN 9789500602532. http://books.google.com.uy/books?id=dTFeuRRZoy0C&pg=PA846&dq=sincitio+neurona&hl=es&ei=rEnlTfWXL8W9tgees9zeCQ&sa=X&oi=book_result&ct=result&resnum=2&ved=0CCoQ6AEwAQ#v=onepage&q=sincitio%20neurona&f=false.
- ↑ Hill, Richard W.; Wyse, Gordon A. (2006). Fisiología Animal. Ed. Médica Panamericana. p. 360. ISBN 9788479039905. http://books.google.com.uy/books?id=HZaC45m9lMMC&pg=PA360&dq=sincitio+Schwann&hl=es&ei=-vziTfrYLaLg0QHCwpmpBw&sa=X&oi=book_result&ct=result&resnum=5&ved=0CDsQ6AEwBA#v=onepage&q=sincitio%20Schwann&f=false.
- ↑ Evans, K.; Stone, A.R. (1977). «A Review of the Distribution and Biology of the Cyst Nematodes Globodera rostochiensis and G. pallida». PANS 23 (2): pp. 178-189.
- ↑ Canto-Sáenz, M.; Mayer De Scurrah, M. (1977). «Races of the potato cyst nematode in the andean region and a new system of classification» (en inglés). Nematologica 23: pp. 340-349. ISSN 0028-2596.
- ↑ Greco, N. (1988). «Potato cyst nematode: Globodera rostochiensis and G. pallida» (en inglés). Nematology 9 (1): pp. 30-35. ISSN 1388-5545.
- ↑ Franco, J.; González, A.; Matos, A.. Manejo Integrado Del Nematodo Quiste de la Papa Globodera Spp. Centro Internacional de la Papa. pp. 46-48. http://books.google.com.uy/books?id=44aVdwvz2pwC&pg=PA46&dq=frecuencia+sincitio&hl=es&ei=Yb4oTv3ME-Tv0gHFxejoCg&sa=X&oi=book_result&ct=result&resnum=5&ved=0CD4Q6AEwBA#v=onepage&q=frecuencia%20sincitio&f=false.
- ↑ Poizo E. (2007). «World distribution of Trichinella spp. infections in animals and humans» (en inglés). Veterinary Parasitology 149 (1-2): pp. 3-21. http://faculty.uml.edu/rhochberg/hochberglab/Courses/Parasite/PDF%20Papers/Nematodes/World%20Distribution%20of%20Trichinella.pdf.
- ↑ Vignau, M. L. (2004). «Triquinosis». Ciencia Hoy 14: pp. 58-65.
- ↑ Aguilar (2002). Tratado de Enfermería Pediátrica Enfermería Mosby. Elsevier España. pp. 692. ISBN 9788481745580. http://books.google.com.uy/books?id=lAX2hkbeBQYC&pg=PA692&dq=sincitio+paramixovirus.&hl=es&ei=gQPLTaLYIdCztweM-rTiBw&sa=X&oi=book_result&ct=result&resnum=4&ved=0CDwQ6AEwAw#v=onepage&q=sincitio%20paramixovirus.&f=false. Consultado el 11 de mayo de 2011.
- ↑ Prats Pastor, Guillermo (2006). Microbiología Clínica. Médica Panamericana. pp. 123. ISBN 9788479039714.
- ↑ Huerta, L et ál (2009). «HIV-Envelope–Dependent Cell-Cell Fusion: Quantitative Studies» (en inglés). The Scientific World Journal 9: pp. 746–763. doi:.
- ↑ Chamizo Pestana, E.G.. UABC. ed. Patología especial y diagnóstico de las enfermedades de los animales domésticos. p. 47. ISBN 9789687326351. http://books.google.com.uy/books?id=tp4MbDTiW_YC&pg=PA47&dq=sincitio+virus+sincitial+respiratorio&hl=es&ei=Sf3bTYKaC4P2tgO9rsXDDg&sa=X&oi=book_result&ct=result&resnum=4&ved=0CD0Q6AEwAw#v=onepage&q&f=false.
- ↑ Shoemaker; Grenvik, Ake (2002). Tratado de medicina crítica y terapia intensiva (4ª, reimpresa edición). Médica Panamericana. pp. 1460-1461. ISBN 9788479035877. http://books.google.com.uy/books?id=d4NfFCXqc2IC&pg=PA1461&dq=origen+t%C3%A9rmino+sincitio&hl=es&ei=8NU0Tvm8G6jY0QHn27DkCw&sa=X&oi=book_result&ct=result&resnum=3&ved=0CDQQ6AEwAg#v=onepage&q&f=false.
Enlaces externos
- Este artículo fue creado a partir de la traducción del artículo Syncytium de la Wikipedia en inglés, concretamente de esta versión, bajo licencia Creative Commons Atribución Compartir Igual 3.0 y GFDL.




Wikimedia foundation. 2010.